Последовательность работ при ремонте: Последовательность ремонта квартиры: Этапы ремонта квартиры
Последовательность работ при ремонте помещений
Существует стандартная последовательность ремонтных работ:
- Вывоз мебели, аппаратуры, вещей и техники (для их хранения можно арендовать небольшой склад или контейнер; при выборе обращайте внимание не только на надежность хранения, но и на температурный и влажностный режим).
- Демонтажные работы с последующим вывозом мусора.
Демонтажные работы
- Покупка и доставка стройматериалов для черновых работ.
- Установка стен и перегородок.
- Электромонтажные работы (без установки светильников и электроприборов).
- Замена сантехнических труб (без установки сантехприборов).
- Прокладка прочих коммуникаций (спутниковое телевидение, интернет, трубы вентиляции и т.д.).
- Выравнивание поверхностей (оштукатуривание, обшивка гипсокартоном). Производится сверху вниз: сначала – потолки, затем – стены и в заключение – пол.
- Финишная шпатлевка.
- Установка новых окон и входной двери (перед началом следующего этапа их следует закрыть пленкой, чтобы избежать загрязнения и механических повреждений).

- Оштукатуривание участков, расположенных рядом с новыми окнами и дверью.
- Грунтовка поверхностей.
- Приобретение материалов для чистовых работ.
- Чистовая отделка потолков (покраска, установка натяжных или навесных потолков). Выполняется по принципу «от дальних углов к входной двери».
- Укладка настенной плитки на кухне и в ванной комнате.
- Укладка напольной плитки.
- Установка сантехприборов. Некоторые специалисты рекомендуют выполнять установку после чистовой отделки стен и укладки полового покрытия, однако, на практике это не слишком удобно. Чтобы избежать повреждений поверхности ванны и душевой кабинки, следует закрыть их пленкой. Как вариант – можно установить только унитаз, а монтаж остальных приборов перенести на более позднее время.
- Чистовая отделка стен
- Укладка полового покрытия.
Укладка полового покрытия
- Монтаж плинтусов (рядом с межкомнатными дверями плинтус оставляем незакрепленным).
- Установка радиаторов.
- Установка межкомнатных дверей.
- Монтаж электроустановочных изделий и осветительных приборов.
- Расстановка мебели, установка и подключение бытовых приборов.
- Декорирование.
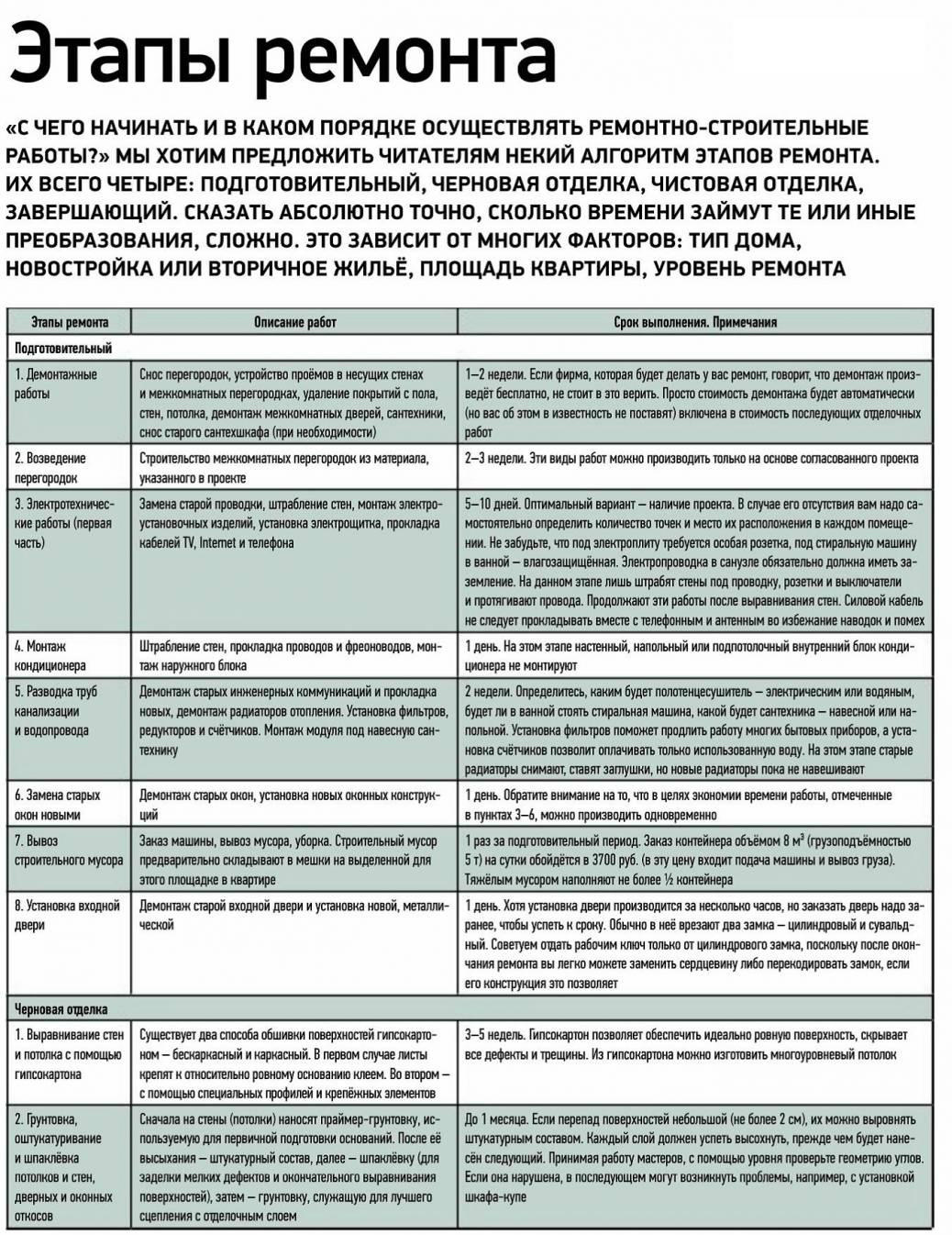
Заключительные этапы ремонта можно несколько варьировать, соблюдая два принципа. Первый – чистовая отделка выполняется сверху вниз и от дальних помещений к выходу. Второй – вначале следует производить «более грязные» работы, а затем – «более чистые».
Чтобы ничего не забыть и не перепутать, выпишите все этапы и подробно расшифруйте каждый пункт. Так вы сможете составить полный перечень всех необходимых работ и определить сумму дополнительных расходов (например, на заказ грузового автомобиля и оплату труда грузчиков).
Расчет количества материалов
Составьте список необходимых материалов на основе предыдущего списка. Измерьте площадь отделываемых поверхностей. Площадь прямой квадратной или прямоугольной поверхности вычислить очень просто. Нужно измерить ширину и высоту и перемножить два этих показателя. Площадь сложных поверхностей рассчитывают, разделяя их на отдельные простые фигуры.
Нужно измерить ширину и высоту и перемножить два этих показателя. Площадь сложных поверхностей рассчитывают, разделяя их на отдельные простые фигуры.
Итак, у вас есть список материалов, вы знаете площадь пола, стен и потолка. Определить объем закупки конкретного материала можно, проконсультировавшись у продавцов специализированного магазина и сообщив ему площадь отделываемой поверхности.
Общедоступные материалы для черновой отделки можно покупать «впритык» или с небольшим «запасом» — в крайнем случае, вы всегда сможете приобрести недостающее количество в любом магазине.
Расчет количества материалов
Плитку, обои, напольные покрытия и другие материалы для чистовой отделки следует приобретать в точном соответствии с рекомендациями продавца (то есть, со значительным «запасом» на случай боя, порчи и т.д.). Не покупайте материал из двух разных партий – возможно несовпадение оттенков. Не приобретайте материал «впритык» — при ошибке в расчетах или случайной порче его может попросту не хватить.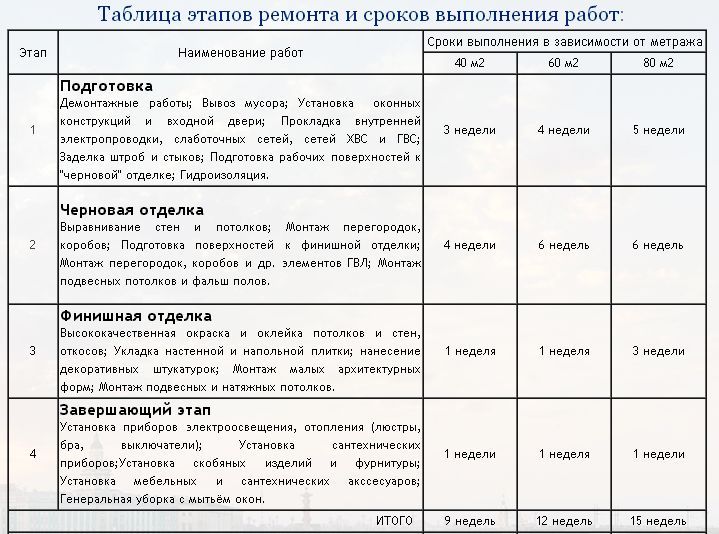 И не забудьте про неизбежные обрезки – тонкие полоски обоев или половинки плитки «до угла» и т.д.
И не забудьте про неизбежные обрезки – тонкие полоски обоев или половинки плитки «до угла» и т.д.
Подсчитав необходимое количество материалов, можно переходить к расчету общей суммы, которая будет потрачена на ремонт. Добавьте к стоимости материалов определенные ранее расходы на прокладку труб и монтаж электроузлов, стоимость светильников, смесителей и сантехнических приборов. Не забудьте про дополнительные расходы (аренда склада для мебели, перевозка вещей, вывоз мусора, доставка стройматериалов, услуги грузчиков). А затем умножьте получившуюся цифру на 1,3-1,5 – примерно такой будет реальная стоимость ремонта.
При ремонте что делать первым – стены или потолок, с чего начать?
Хорош тот ремонт, что запланирован от и до. И речь идет даже не о дизайн-проекте, а о последовательности действий. Представьте, что у вас уже закуплены материалы, сделан необходимый демонтаж. Что при ремонте делать первым – потолок или стены? Или вовсе пол? Раньше ответ звучал однозначно: работы ведутся сверху вниз. Теперь же, при наличии разных отделочных материалов, возможны варианты. Давайте посмотрим, как практичнее выстроить последовательность работ.
Теперь же, при наличии разных отделочных материалов, возможны варианты. Давайте посмотрим, как практичнее выстроить последовательность работ.
Что первое – потолок или стены – при обычном ремонте?
Любой строитель вам подтвердит, что по всем правилам ремонт делается, начиная с потолка, затем стены и в последнюю очередь – пол. Подразумевается, что обломки штукатурки, могут повредить новое напольное покрытие, а пыль при работах с потолком осядет на стенах. Какие работы под этим подразумеваются?
- Демонтаж старых покрытий – удаление обоев, старых покрытий потолка.
- Зачистка поверхностей – вся пыльная работа.
- Если необходимо выравнивание, то сначала выравнивают потолок, потом стены.
- Покраска потолка.
- Поклейка обоев или нанесение штукатурки, покраска стен.
Если потолок не красить, а наклеивать панели или вовсе обои (да, так тоже делают) – как в таком случае? Точно так же. При этом потолочный плинтус лучше приклеивать уже после обоев – он должен скрывать стык.
При этом потолочный плинтус лучше приклеивать уже после обоев – он должен скрывать стык.
Важно: и потолочные панели, и плинтус, и декоративные бесцветные накладки вокруг люстр мастера-отделочники рекомендуют сверху покрасить в один слой обычной краской для потолков. Так элементы будут выглядеть естественнее.
Нестандартные покрытия: что сперва потолок или стены?
Теперь давайте поговорим о различных нестандартных покрытиях – точнее, современных, которые используются все чаще при дизайнерском ремонте. Это:
- обои из ткани;
- облицовочные панели – из камня, дерева, МДФ, ПВХ;
- натяжные и подвесные конструкции – для потолка и стен;
- кессоны – деревянные, из полиуретана;
- фальшбалки и многое другое.
Чаще всего вопросы возникают именно по натяжным потолкам. Дело в том, что они могут монтироваться и вне ремонта, то есть, стены и пол остаются нетронутыми: часть стены просто уходит за грань поверхности, вместе с обоями.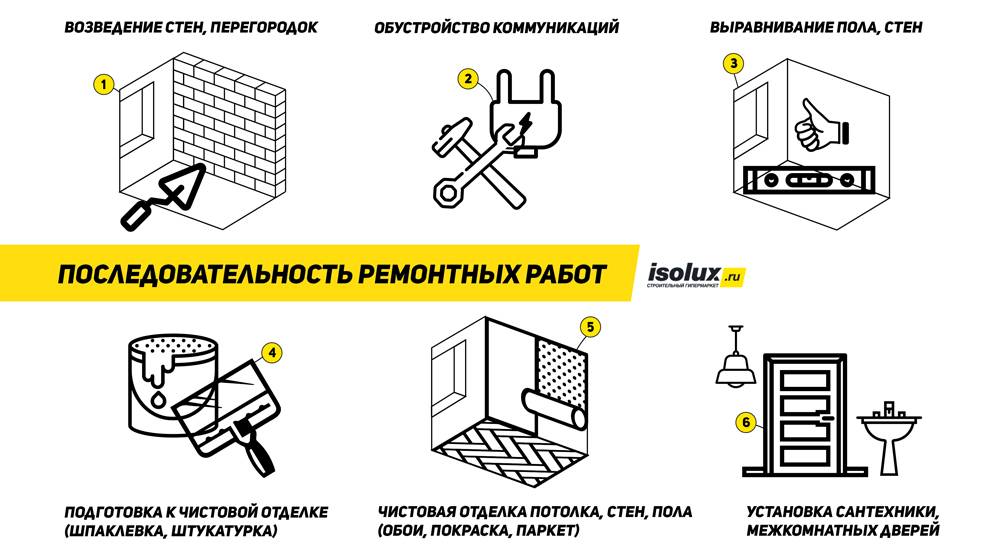 Значит ли это, что при ремонте нужно сначала все же клеить обои? Мастера имеют разные мнения на этот счет.
Значит ли это, что при ремонте нужно сначала все же клеить обои? Мастера имеют разные мнения на этот счет.
Особенности национального ремонта: распространенные ошибки
Стены – вторыми. Это основной постулат, опирающийся на неизменное появление пыли при монтаже. Даже если вы ставите простой одноуровневое натяжное полотно, то в любом случае придется сверлить стены под крепления, а это пыль. С навесными конструкциями это тоже актуально: если подразумеваются крепления не только к стенам, но и к потолку.
Стены – первыми. Тот редкий случай, когда проще пропылесосить поверхность или воспользоваться щеткой, чем спрятать границы. Возникает при монтаже многоуровневых конструкций – к коробам из гипсокартона сложно подобрать плинтус, поэтому стыкможет быть открытым. Если у вас тканевые обои, имеет смысл их довести до верха, и поверх уже устанавливать короб. Та же история со штукатуркой и жидкими обоями. Но все же лучше сначала сделать потолок.
Тот редкий случай, когда проще пропылесосить поверхность или воспользоваться щеткой, чем спрятать границы. Возникает при монтаже многоуровневых конструкций – к коробам из гипсокартона сложно подобрать плинтус, поэтому стыкможет быть открытым. Если у вас тканевые обои, имеет смысл их довести до верха, и поверх уже устанавливать короб. Та же история со штукатуркой и жидкими обоями. Но все же лучше сначала сделать потолок.
И если с кессонами все понятно – они устанавливаются по принципу потолочной плитки, – то ситуация с фальшбалками может быть не всегда очевидной. Они могут крепиться не только к потолку, но и упираться непосредственно в стены и прикручиваться там (чтобы был доступ к коммуникациям внутри, если речь о полых конструкциях). При штукатурке их можно обойти, а если у вас обои, то можно сделать так: сначала вчистую сделать потолок, затем наклеить обои и уже потом прикрепить балку.
Если в вашем интерьере подразумеваются натяжные покрытия и для стен, то их должны устанавливать мастера в порядке, который подразумевает конструкция.

А, может, сначала пол?
Действительно, такой вопрос возникает, когда с полом планируется делать что-то существенное – например, обновление стяжки. Размешивание и заливка бетона поверх керамзита – пыльное дело. Причем, в некоторых случаях – например, при ремонте в старом фонде, – после демонтажа пола не всегда получится попросту поставить стремянку – некуда, да и перекрытия будут мешаться.
Что делать в этом случае? Примерный порядок действий такой:
- демонтаж – потолок (при необходимости – до перекрытий), затем стены, потом пол;
- тщательное обеспыливание и уборка вообще всего строительного мусора;
- обустройство стяжки и заливка пола;
- выравнивание стен и потолка;
- отделка потолка;
- наклейка обоев или штукатурка;
- укладка напольного покрытия.
Таким образом, вся черновая работа делается в первую очередь, а декоративная отделка наносится в порядке сверху вниз.

Главное при любой последовательности ремонта – тщательное обеспыливание поверхностей. Чем масштабнее ремонт, тем более серьезные средства пригодятся. Например, при капитальном демонтаже не обойтись без строительного пылесоса, а если вы делаете легкую «косметику», то хватит обычного бытового, щетки и ведра с тряпкой.
5 советов, как сэкономить на ремонте квартиры
Как и на чем можно сэкономить деньги при ремонте квартиры: полезные советы
Мы рассказали вам о различных вариантах последовательности ремонта, в частности, о том, что делать вперед – потолок или стены. Несмотря на то, что существует некая традиционная последовательность, всегда стоит учитывать особенности используемых материалов и масштаба ремонта.
Последовательность ремонта квартиры. Ремонтные работы в новостройке
Подготовка к ремонту
Сразу браться за работу нельзя, сначала нужно все продумать. Какие-то стены нужно снести, где-то наоборот возвести новые перегородки. Там будет висеть телевизор, там кондиционер, вы должны представить себе будущую квартиру, не до мелочей, конечно, хотя бы в общих чертах.
Для начала каких-либо работ следует организовать рабочее пространство, вам понадобится временная сантехника и электричество. Также нужно закупить необходимые инструменты.
Демонтажные работы
А вот теперь можно начинать ломать и крушить то, что вам не нравится. Сносим ненужные (не несущие) стены.
Старые же окна и двери пока лучше оставить, без них далеко не пойдешь, а новые жалко пачкать или хуже того — наносить всевозможные повреждения.
Перепланировка
Если речь идет о последовательности ремонта в новостройке, то в последнее время все чаще квартиры продаются с учетом того, что все хотят не только индивидуальный дизайн, но еще и планировку. Выглядят такие квартиры, как огромная коробка, и где вы хотите сделать перегородку — это уже ваше личное дело.
Так вот на этом этапе самое время воздвигать новые стены. Они могут быть как кирпичные, так и гипсокартонные, тут вопрос только в прочности и звукоизоляции.
Разводка коммуникаций
Теперь уже нужно четко продумать что и где будет находиться, и главное ничего не забыть! Все необходимые розетки, телевидение, телефон, кондиционер, акустические системы, интернет. Также не забываем про отопление, водопровод, канализацию.
Важно! Тщательно продумайте все эти моменты, потому что потом исправить что-то будет либо очень сложно, либо невозможно.
Замена окон и дверей
Сейчас самое время поменять окна, а заодно и входные двери. Так как цементные работы будут включать в себя штукатурку откосов и углов, после которых что-либо заменять уже будет поздно.
Так как цементные работы будут включать в себя штукатурку откосов и углов, после которых что-либо заменять уже будет поздно.
Подготовка поверхностей — цементные работы
Все имеющиеся поверхности, а именно — пол, потолок, стены, нужно выровнять.
Начинаем с штукатурки потолка, при необхожимости, потом выравниваем стены (не обшиваем гипсокартоном, а именно, штукатурим), и, наконец, приходим к стяжке пола.
Важно! Если в вашей квартире планировались теплые полы, то укладывать их нужно до стяжки пола.
Обшивка гипсокартоном
Выравнивание стен гипсокартоном, монтаж полок и потолков из гипсокартона проводятся уже после высыхания стяжки и штукатурки, так как тут повышенная влажность будет вашим злейшим врагом.
Шпатлевание стен и потолков
Перед малярными работами, нужно зашпаклевать поверхности, при чем более тщательная шпаклевка потребуется при покраске, так как если обои еще могут скрыть какие-то мелкие дефекты, то краска только выставит их напоказ.
Облицовка поверхностей
Укладка плитки на стены и пол
С чего начать укладывать плитку с пола или стен — вопрос спорный, некоторые начинают с пола, что логично, но кафель сильно испачкается, когда вы перейдете к стенам. Некоторые же придумали весьма хитрый ход — они начинают со стен, но не укладывают самый нижний ряд, а оставляют там зазор высотой в одну плитку плюс будущая плитка пола, фиксируя плитку на стене деревянной рейкой.
Позже, когда пол уже уложен, они просто убирают рейку и аккуратно укладывают нижний ряд.
После укладки производится затирка швов.
Инфо! Когда плитка уложена можно устанавливать сантехнику.
Поклейка обоев и покраска стен и потолков
Начинать следует с потолков (вместо покраски и поклейки обоев можно использовать потолочную плитку). Обои или краска — это ваш выбор, каждый материал имеет свои преимущества и недостатки. Также не забывайте об обоях под покраску.
Что же касается выбора вида краски и выбора обоев, то здесь отталкиваться следует от особенностей помещения и вашей дизайнерской задумки.
Помимо обоев и краски существует множество не менее качественных и интересных материалов, например, декоративная штукатурка, панели, пробка и многое другое, подробнее читайте статью Чем отделать стены в квартире.
Укладка напольного покрытия
Плитку нужно было уложить раньше, а остальные покрытия (ламинат, паркет, линолеум) при монтаже не производят много грязи, поэтому и укладываются в конце. О существующих покрытиях для пола и о том как их выбрать читайте на нашем сайте.
Установка межкомнатных дверей и плинтусов
Пол есть, стены есть, осталось поставить двери и установить плинтуса. После этого можно устанавливать кухонную и другую мебель, ну и открывать шампанское.
Последовательность и список работ при ремонте квартиры подробно расписана в таблице.
Удачного ремонта!
Больше информации
Задать вопрос
Последовательность ремонта квартиры своими руками с нуля: очередность работ
Рано или поздно наступает момент, когда нужно делать в квартире ремонт. Если у вас есть средства, вы можете нанять специалистов, которые сделают все как надо. Но как быть, когда такой возможности нет, и ремонтировать квартиру придется самостоятельно? Ведь невозможно сделать хороший ремонт, ничего в этом не понимая.
Если у вас есть средства, вы можете нанять специалистов, которые сделают все как надо. Но как быть, когда такой возможности нет, и ремонтировать квартиру придется самостоятельно? Ведь невозможно сделать хороший ремонт, ничего в этом не понимая.
Поэтому, перед тем как начинать все ломать и рушить, необходимо знать, что и в каком порядке нужно делать. При ремонте квартиры грамотно составленная последовательность работ поможет вам сэкономить не только время, но и средства.
Как быть, если нет возможности съехать с квартиры на время ремонта
Квартиру желательно ремонтировать всю полностью и одновременно, чтобы грязь не таскалась из одной комнаты в другую. Но не стоит переживать, если по какой-то причине вы не можете съехать на время проведения ремонтных работ.
В таком случае начинать ремонт следует с помещения, которое находится дальше всех от входной двери, и заканчивать его в коридоре.
Следуя этому простому правилу, вам не придется по только что отремонтированным комнатам носить строительный материал и различный мусор. Тем самым вы оградите себя от неприятных сюрпризов, которые могу случиться во время ремонтных работ в вашем доме. И если вам предстоит делать ремонт в квартире, не выезжая из нее, строго придерживайтесь последовательности намеченных вами действий.
Тем самым вы оградите себя от неприятных сюрпризов, которые могу случиться во время ремонтных работ в вашем доме. И если вам предстоит делать ремонт в квартире, не выезжая из нее, строго придерживайтесь последовательности намеченных вами действий.
Как делать ремонт в квартире
Итак, вы определились с объемом работ. Строительные материалы уже закуплены. Мебель вынесли. Теперь можно приступать.
Начальный этап ремонтных работ самый грязный и самый трудоемкий. Первым делом нужно удалить с потолка и стен старые обои, краску и штукатурку. А если вы запланировали замену старых окон и дверей или перепланировку комнат, то это необходимо делать в первую очередь. Процесс демонтажа всегда сопровождается большим количеством грязи и пыли. С него и нужно начинать любой ремонт, чтобы в дальнейшем при проведении отделочных работ ничто не испортило покрытие стен и потолков.
Затем следует уделить внимание электропроводке, водопроводным трубам и системе отопления. Если требуется их замена, то это следует сделать до того, как вы приступите к отделке стен.
Последовательность ремонта квартиры должна предусматривать выполнение самых грязных работ на начальном этапе. А замена электропроводки, установка добавочных розеток и выключателей, газосварочные работы обязательно станут причиной появления дополнительного мусора и грязи.
После того как вы удалили старую отделку, установили новые окна и заменили все коммуникации, можно приступать к выравниванию стен и потолков.
На этом этапе ремонтных работ устанавливаете гипсокартонные конструкции, грунтуете и шпаклюете поверхности. Ровняя стеновые перегородки, не забудьте откорректировать места прилегания плинтусов.
Закончив со стенами, вам следует заняться полом. Во время капитального ремонта квартиры последовательность работ такова, что, только придав половому покрытию ровную поверхность, вы можете устанавливать межкомнатные двери. Но это не значит, что нужно сразу покрывать его краской, ламинатом или линолеумом. Это вы сделаете на завершающем этапе ремонтных работ.
Если в вашей квартире деревянное половое покрытие, и вы не собираетесь его менять, то перед тем, как устанавливать новые двери, его желательно обработать электрорубанком или специальной шлифовальной машиной.
Незначительные неровности бетонного пола устраняются с помощью специальной ремонтной смеси, которую можно приобрести в строительном магазине.
Ровнять пол следует до того, как вы начнете отделывать потолки и стены.
И нарушать эту последовательность ремонта квартиры не стоит. В процессе выравнивания полового покрытия образуется огромное количество пыли. Если к этому моменту потолок и стены в вашей квартире будут уже оклеены обоями и покрыты краской, они, как магнит, притянут к себе все микрочастицы и будут иметь непрезентабельный вид.
Поэтому, прежде чем приступать к отделочным работам, необходимо во всей квартире выровнять пол.
Закончив выравнивать половую поверхность и устанавливать двери, можно начинать класть плитку, красить потолки и наклеивать обои. Финальным аккордом в вашем ремонте будет отделка полов. Независимо от того, как вы хотите это сделать — покрыть паркетом, ламинатом, линолеумом или краской — делать это нужно в последнюю очередь. Так как все уже окрашено и приклеено, ничто не испортит ваше половое покрытие.
Финальным аккордом в вашем ремонте будет отделка полов. Независимо от того, как вы хотите это сделать — покрыть паркетом, ламинатом, линолеумом или краской — делать это нужно в последнюю очередь. Так как все уже окрашено и приклеено, ничто не испортит ваше половое покрытие.
В принципе, нет ничего сложного в том, чтобы своими руками сделать ремонт квартиры, если следовать такой последовательности ремонтных работ. Всю выше изложенную информацию можно отобразить в простом пошаговом алгоритме:
- Демонтаж старой отделки;
- Перепланировка;
- Монтаж новых окон;
- Замена электропроводки, водопроводных труб, системы отопления;
- Установка гипсокартонных конструкций;
- Шпаклевка стен и потолков;
- Выравнивание пола;
- Установка межкомнатных дверей;
- Отделочные работы;
- Покрытие полов.
Планируя ремонт квартиры, составьте для себя подобный план с указанием вида работ и сроком их выполнения.
 Это поможет вам не только сделать все грамотно, но и сэкономить время.
Это поможет вам не только сделать все грамотно, но и сэкономить время.Даже если вы решите воспользоваться услугами специалистов, то составленная вами последовательность ремонта квартиры, позволит контролировать ход их действий. Недаром же говорят: «Доверяй, но проверяй».
Ремонтные работы в новостройке
Если вы стали счастливым обладателем квартиры в только что построенном доме, то, прежде чем въехать в нее, вам нужно будет делать там ремонт с нуля. Обычно любое жилище в новостройках представляет собой бетонную коробку с окнами, центральными канализацией и водопроводом. Все остальное вам придется делать самостоятельно.
Можно обратиться за помощью к специалистам. Но если ваши финансы не позволяют сделать это, то паниковать не стоит, вы способны самостоятельно отремонтировать ваше новое жилье.
Последовательность ремонта квартиры в новостройке немного отличается от порядка работ в доме, где жили много лет. Вся разница в том, что вам не нужно сглаживать изъяны стареющих поверхностей, удалять обветшалую внутреннюю отделку помещения, менять старые трубы и электропроводку. Вы сразу будете устанавливать новый материал.
Вы сразу будете устанавливать новый материал.
Начните с проведения проводки, монтажа розеток и выключателей. Об этом нужно позаботиться в первую очередь.
Бывали случаи, когда новоселы, торопясь въехать в новую квартиру, быстро отделывали помещения, совершенно забыв провести в них электричество. А когда они вспоминали об этом, было уже поздно: потолки побелены, стены оклеены. И приходилось все начинать сначала. Чтобы не попасть в подобную ситуацию, делая в квартире ремонт с нуля, не пренебрегайте последовательностью выполнения ремонтных работ.
Закончив монтаж электропроводки, установите сантехнику, подведите к ней воду и начинайте ровнять потолки и стены. Некоторые люди ошибочно полагают, что в наш век высоких технологий поверхности в новостройках не требуют выравнивания. К сожалению, их нельзя назвать идеально ровными, и вам никак не избежать этого этапа ремонтных работ.
Завершив ровнять стены и потолки, займитесь полом. Для того чтобы постелить паркет, ламинат или линолеум, хотите вы или нет, но вам придется выравнивать половую поверхность с помощью бетонной стяжки. Смеси для такого вида работ продаются в строительных магазинах, и вам не составит особого труда приготовить раствор и залить им пол.
Смеси для такого вида работ продаются в строительных магазинах, и вам не составит особого труда приготовить раствор и залить им пол.
Когда половая стяжка высохнет, можете смело приступать к отделочным работам. Но помните, что и в новой квартире последовательность ремонта предусматривает установку межкомнатных дверей и укладку полового покрытия только тогда, когда все уже окрашено и оклеено.
Мы надеемся, что наши советы и рекомендации помогут вам грамотно распланировать и выполнить все этапы ремонтных работ в вашем доме.
Что такое заказ на работу? 6 шагов для идеального заказа на работу
Заказы на работу — это двигатель вашей операции по техническому обслуживанию. Они придают силу вашей команде и перемещают работу из точки А в точку Б. Но в мире миллионы двигателей, от ржавых лодок до мощных шпилек. Эта статья посвящена освоению порядка работ по техническому обслуживанию, чтобы ваша операция работала так же гладко, как роскошный спортивный автомобиль.
Шаблон для создания идеального рабочего задания каждый раз (не проводя часы за рабочим столом)
Содержание
Что такое заказ на работу?
Заказ на работу — это документ, в котором содержится вся информация о задаче обслуживания и описывается процесс выполнения этой задачи.Заказы на работу могут включать сведения о том, кто санкционировал задание, объем, кому оно назначено и что ожидается.
Заказы на работу — это двигатель вашего технического обслуживания. Они придают сил вашей команде и перемещают работу из точки А в точку Б.
Заказы на работу имеют решающее значение для технического обслуживания организации. Они помогают всем, от менеджеров по техническому обслуживанию до техников, организовывать, назначать, расставлять приоритеты, отслеживать и выполнять ключевые задачи. При правильном выполнении рабочие задания позволяют собирать информацию, делиться ею и использовать ее для максимально эффективного выполнения работы.
Несмотря на то, что рабочее задание и рабочий запрос звучат похоже, у них есть несколько ключевых отличий. Запрос на работу используется не обслуживающим персоналом, чтобы информировать команду обслуживания о задаче. Например, оператор станка может подать заявку на работу, когда оборудование выходит из строя. Запрос на работу рассматривается менеджером по обслуживанию, который добавляет дополнительную информацию, планирует задачу и назначает ее техническому специалисту. Запрос на работу теперь является заказом на работу.
Как создать идеальное рабочее задание за шесть шагов
Создайте идеальный заказ на работу
Процесс заказа на работу
Жизненный цикл каждого заказа на техническое обслуживание состоит из трех основных этапов: создание, выполнение и запись.Эти этапы можно разбить на несколько этапов. Понимание каждого шага и наличие надежного процесса заказа на работу гарантирует, что задачи не застревают на одной фазе и не превращаются в невыполненную работу.
Шаг №1: Задача идентифицирована
Задачи обслуживания делятся на две группы: плановое обслуживание и внеплановое обслуживание. Плановое техническое обслуживание включает в себя все работы, о которых вы знаете заранее, например плановые проверки, а незапланированное обслуживание включает все задачи, которые вы не можете предвидеть, например неожиданную поломку.
Шаг № 2: создается запрос на обслуживание
Детали задания собраны и переданы команде техобслуживания для дальнейших действий. Например, когда машина выходит из строя, оператор создает запрос на работу и отправляет его в техническое обслуживание. Если задача запланирована, заказ на работу создается и запускается в нужное время.
Шаг № 3. Рабочее задание имеет приоритет и запланировано
Некоторые задания требуют больше времени, чем другие.Перегоревшую лампочку не нужно чинить немедленно, но сломанную конвейерную ленту можно.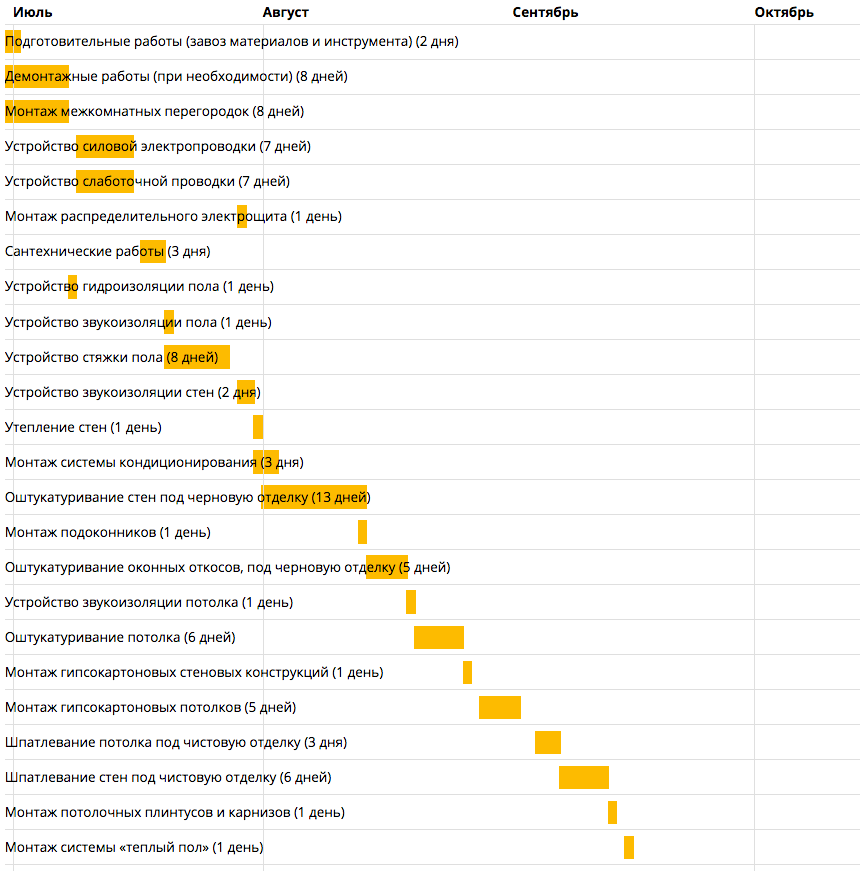 Вот почему вам нужно расставлять приоритеты для каждого рабочего задания, которое попадает на ваш стол.
Вот почему вам нужно расставлять приоритеты для каждого рабочего задания, которое попадает на ваш стол.
После расстановки приоритетов пришло время составить расписание. Заказы на работу можно планировать на основе установленного срока, запланированных триггеров обслуживания или выделенных блоков времени. Установление крайнего срока позволяет всем быть подотчетными и информированными, чтобы ничего не упустить.
Шаг № 4: Работа назначена и завершена
Пришло время превратить слова на странице в действие.Заказ на работу назначается техническому специалисту, который выполняет задачу. Это может быть пятиминутная проверка оборудования, а может быть сложный ремонт, занимающий несколько дней.
Шаг № 5: Заказ на работу закрывается и документируется
После выполнения всех условий заказа на работу его можно закрыть. Руководителям может потребоваться подписать наряд на работу для соответствия требованиям. После закрытия заказ на работу сохраняется. Правильно организованный журнал рабочих заданий имеет решающее значение для создания истории активов, анализа прошлых решений, подготовки к аудиту и многого другого.
Правильно организованный журнал рабочих заданий имеет решающее значение для создания истории активов, анализа прошлых решений, подготовки к аудиту и многого другого.
Шаг № 6: Рабочий заказ анализируется и/или перерабатывается
Закрытые рабочие задания содержат ценную информацию. Они могут дать представление о ваших процессах и системах, которые можно использовать для точной настройки вашей работы. Наличие журнала рабочих заданий также позволяет техническим специалистам быстро обнаруживать любые пропущенные шаги или альтернативные решения, если проблема возникает снова.
Что должно быть в наряде на работу?
Заказы на работу похожи на все, что производит ваше предприятие — они должны быть выполнены качественно и без дефектов.Если одна часть процесса отключена, это может повлиять на всю линию. Итак, какая информация составляет отличный заказ на работу?
- Актив : Какая часть оборудования нуждается в ремонте?
- Описание проблемы : В чем проблема? Что вы слышали, видели, обоняли или чувствовали в момент неудачи или перед ней?
- Объем работ : Какая работа требуется для выполнения работы? Какие навыки необходимы?
- Необходимые детали и инструменты : Есть ли какие-либо детали, которые необходимо заменить, или какие-либо специальные инструменты, которые необходимо использовать?
- Примечания по охране труда и технике безопасности : Какие процедуры безопасности и оборудование необходимы? Были ли какие-либо несчастные случаи или промахи при работе над аналогичной проблемой или активом?
- Дата запроса : Когда было создано и отправлено рабочее задание?
- Имя/отдел/контактное лицо заявителя : Кто создал и отправил заказ на работу?
- Ожидаемая дата завершения : Когда должен быть выполнен этот рабочий заказ?
- Фактическая дата завершения : Когда заказ на работу был завершен и закрыт?
- Ожидаемое количество часов работы : Сколько часов потребуется для выполнения заказа на работу?
- Фактические часы работы : Сколько часов потребовалось для выполнения заказа на работу?
- Контрольный список задач : Существует ли пошаговое руководство по выполнению необходимой работы?
- Приоритет : Насколько важен этот заказ на работу? Высокий, средний или низкий?
- Назначен : Кто будет выполнять работу? Требуется ли более одного человека? Требуется ли внешний подрядчик?
- Связанные документы : Существуют ли ресурсы, которые могут помочь более эффективно выполнить заказ на работу, такие как СОП, руководства, диаграммы, видео, история активов, заказы на покупку или изображения?
- Примечания : Есть ли какие-либо другие наблюдения, которые могут быть полезны при выполнении заказа на работу или просмотре заказа на работу после его закрытия, например, частота возникновения проблемы, методы устранения неполадок или найденное решение?
Шаблон для создания контрольных списков обслуживания, которые работают каждый раз
Получить шаблон
Управление заказами на техническое обслуживание
Простое создание отличного заказа на работу не гарантирует успеха. Этот рабочий заказ также должен управляться должным образом. Надежная система управления жизненным циклом заказа на работу обеспечивает плавный переход от одного шага к другому. Это поможет вам избежать всевозможных проблем, таких как отсутствие подотчетности, высокие затраты, увеличение времени простоя и сокрушительное отставание. Давайте посмотрим на плюсы и минусы нескольких систем управления рабочими заданиями и на то, как они оцениваются.
Этот рабочий заказ также должен управляться должным образом. Надежная система управления жизненным циклом заказа на работу обеспечивает плавный переход от одного шага к другому. Это поможет вам избежать всевозможных проблем, таких как отсутствие подотчетности, высокие затраты, увеличение времени простоя и сокрушительное отставание. Давайте посмотрим на плюсы и минусы нескольких систем управления рабочими заданиями и на то, как они оцениваются.
Посмотрите, как выглядит упрощенное управление обслуживанием
Попробуйте бесплатно
Ручка и бумага
Рабочие задания управлялись ручкой и бумагой со дня их изобретения.Письменные заказы на работу экономичны и знакомы. Бумага — это инструмент, которым удобно пользоваться всем. Это практически не требует обучения, первоначальные затраты довольно низкие, и есть бумажный след, когда вам нужно проверить прошлую работу.
Однако у этой системы есть серьезные недостатки. Бумажные файлы легко ошибаются, теряются или повреждаются. Они громоздки и требуют времени, чтобы найти, извлечь и отсортировать. Неточная информация с большей вероятностью попадет в заказ на работу, поскольку детали часто записываются после инцидента.Время отклика на рабочие запросы также медленнее. В совокупности эти факторы делают работу менее эффективной и могут стоить вам больших денег в будущем.
Они громоздки и требуют времени, чтобы найти, извлечь и отсортировать. Неточная информация с большей вероятностью попадет в заказ на работу, поскольку детали часто записываются после инцидента.Время отклика на рабочие запросы также медленнее. В совокупности эти факторы делают работу менее эффективной и могут стоить вам больших денег в будущем.
Некоторые задания требуют больше времени, чем другие. Перегоревшую лампочку не нужно чинить немедленно, но сломанную конвейерную ленту можно. Вот почему вам нужно расставлять приоритеты для каждого рабочего задания, которое попадает на ваш стол.
Белые доски
Белые доски— еще один старый резерв для отделов технического обслуживания. Стоимость материалов не слишком увеличивает бюджет, и, безусловно, легко иметь все рабочие задания, доступные для просмотра и обновления в одном централизованном месте.
Подобно ручке и бумаге, белые доски имеют ряд серьезных ограничений. Ведение записей — это огромная головная боль, и чрезвычайно сложно извлечь информацию из любых записей, которые вам действительно удается получить. Это делает почти невозможным создание истории активов, подготовку к аудиту и создание отчетов о рабочих заданиях. Процесс управления рабочими заданиями также тормозится, поскольку операторам и техническим специалистам необходимо обращаться в центральное место для подачи или просмотра рабочих запросов.
Это делает почти невозможным создание истории активов, подготовку к аудиту и создание отчетов о рабочих заданиях. Процесс управления рабочими заданиями также тормозится, поскольку операторам и техническим специалистам необходимо обращаться в центральное место для подачи или просмотра рабочих запросов.
Электронные таблицы Excel
Электронные таблицы Excel— это шаг вперед по сравнению с ручкой, бумагой и белой доской.Это делает записи цифровыми, поэтому файлы с меньшей вероятностью будут повреждены или потеряны. Также проще искать информацию и создавать отчеты, используя эту информацию.
Но хотя электронные таблицы немного поднимают планку, есть некоторые факторы, которые делают их шаткой основой для управления заказами на техническое обслуживание. Некоторые электронные таблицы привязаны к отдельным компьютерам, что затрудняет просмотр актуальной информации о рабочем задании. Даже если они основаны на облаке, электронные таблицы не имеют возможности автоматически запускать рабочие задания, что чрезвычайно затрудняет профилактическое обслуживание. Ввод данных и создание отчетов требуют длительного времени работы за компьютером и ноу-хау. Существует также ограниченная возможность отслеживать ход выполнения заказов на работу, что оставляет вас на шаг позади.
Ввод данных и создание отчетов требуют длительного времени работы за компьютером и ноу-хау. Существует также ограниченная возможность отслеживать ход выполнения заказов на работу, что оставляет вас на шаг позади.
Программное обеспечение CMMS
Одним из самых больших преимуществ компьютеризированных систем управления техническим обслуживанием (CMMS) является использование в них мобильных и облачных технологий. Такое программное обеспечение для заказа работ по техническому обслуживанию позволяет всем специалистам по техническому обслуживанию создавать, отслеживать, выполнять и анализировать задачи в режиме реального времени из любого места — будь то место аварии или пляж на Гавайях.Технические специалисты могут брать с собой рабочие задания, истории активов, документы и изображения, куда бы они ни направлялись. Они также уведомляются о новых заказах на работу, как только они отправляются или инициируются. Отчеты извлекают данные из заказов на техническое обслуживание по стоимости, эффективности и другим показателям. Для тех, кто не занимается техническим обслуживанием, отправка запроса на работу через CMMS может дать им большее чувство ответственности за эту работу. Они могут отслеживать статус своих запросов, и это устраняет дублирование рабочих заданий. Это ключевой способ увеличить TPM на вашем объекте и уменьшить потребность в получении обновлений или разъяснений по задаче.
Для тех, кто не занимается техническим обслуживанием, отправка запроса на работу через CMMS может дать им большее чувство ответственности за эту работу. Они могут отслеживать статус своих запросов, и это устраняет дублирование рабочих заданий. Это ключевой способ увеличить TPM на вашем объекте и уменьшить потребность в получении обновлений или разъяснений по задаче.
Хотя за программным обеспечением CMMS будущее, оно требует более высоких начальных цен, требует исключительного обучения и культуры, чтобы сделать систему успешной, и часто требует более продвинутых методов обслуживания. Хотя программное обеспечение CMMS имеет свои недостатки, долгосрочные преимущества системы более чем компенсируют любые первоначальные недостатки.
Суть
Заказы на работу — основа отличного обслуживания. При правильном управлении они придают вашей команде стабильность и структуру, необходимые для эффективной работы.Хорошо составленный заказ на техническое обслуживание и процесс заказа на работу упрощают создание программы профилактического обслуживания и реагирование на внеплановое обслуживание.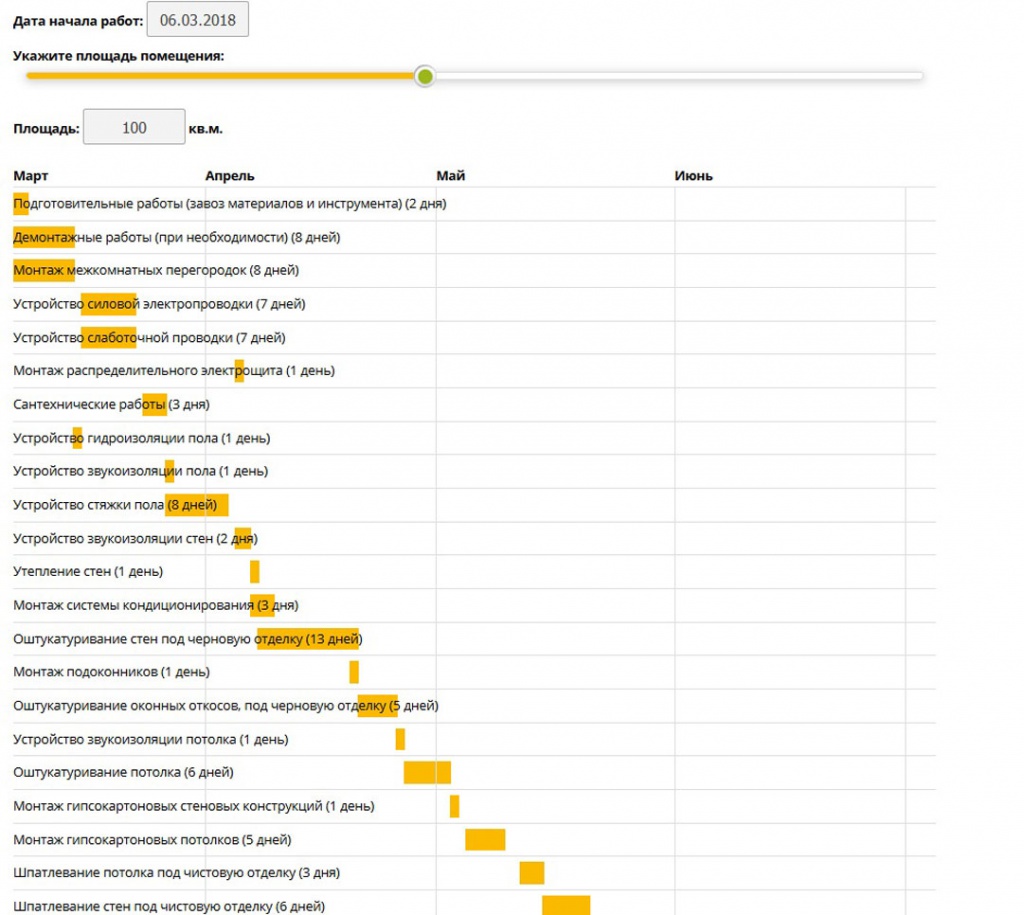 Роли определены, рабочие процессы более плавные, задачи отслеживаются, а информация хорошо документируется. Выбор правильных инструментов и систем для управления рабочими заданиями — решающая заключительная часть головоломки. Когда все это складывается вместе, ваша компания может освоить основы технического обслуживания и искать новые пути роста и успеха.
Роли определены, рабочие процессы более плавные, задачи отслеживаются, а информация хорошо документируется. Выбор правильных инструментов и систем для управления рабочими заданиями — решающая заключительная часть головоломки. Когда все это складывается вместе, ваша компания может освоить основы технического обслуживания и искать новые пути роста и успеха.
Полный 7-этапный процесс укладки асфальтового покрытия
Никогда не поздно узнать что-то новое, и когда дело доходит до укладки асфальтового покрытия, есть много вещей, о которых домовладельцы, владельцы коммерческой недвижимости и муниципальные служащие могут не знать .
В компании Wolf Paving мы являемся подрядчиками по укладке асфальта из Милуоки, Окономовок и Мэдисон, которые гордятся тем, что обучают людей всему, что касается асфальта — это наша страсть.
Взгляните, пожалуйста, на этот пошаговый обзор того, что требуется для укладки асфальтового покрытия — от сноса до получения гладкой новой поверхности.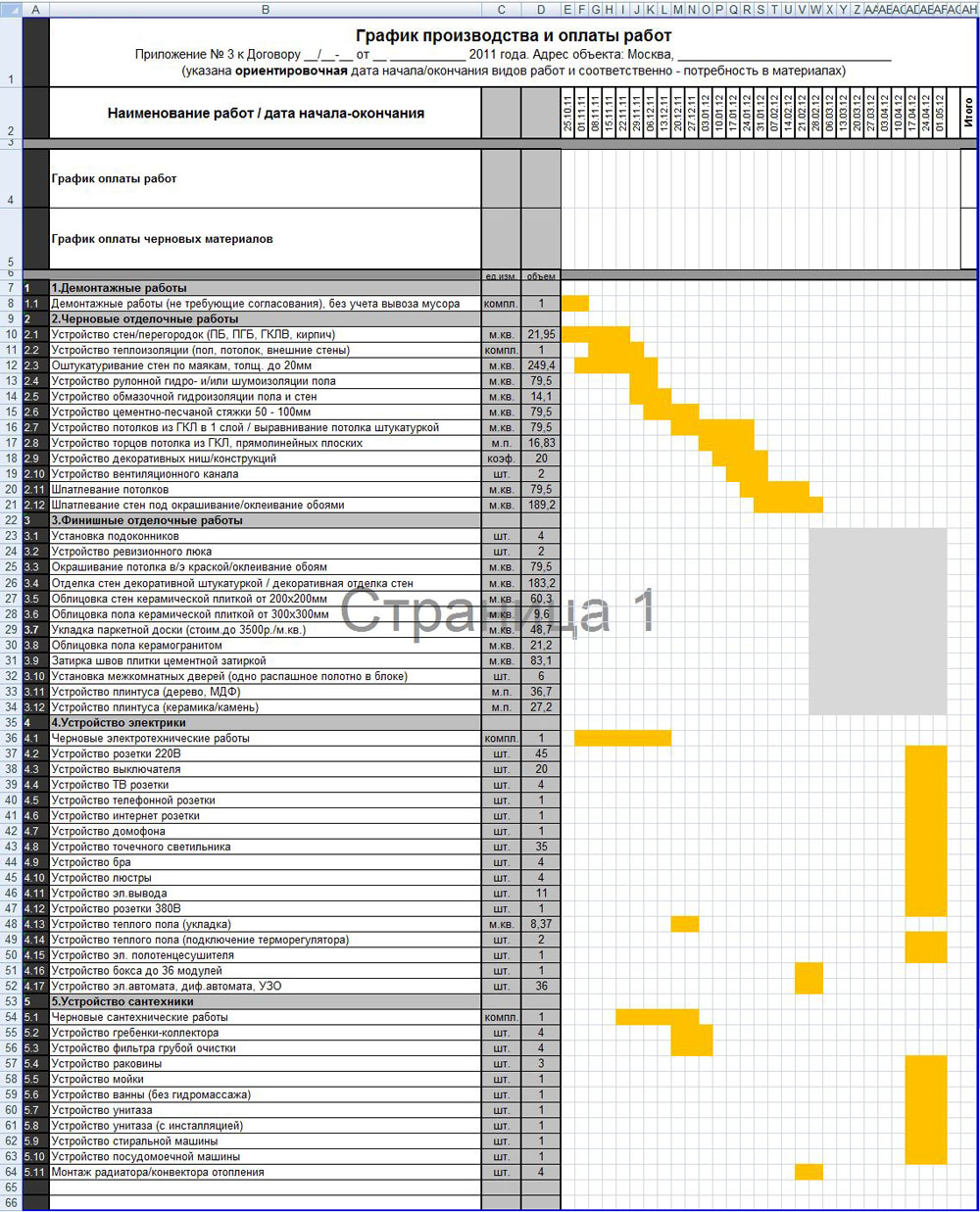
Первым шагом в процессе укладки асфальта является удаление существующего покрытия, будь то асфальт, бетон или брусчатка.
- Снос и вывоз выполняются с использованием тяжелой техники, включая небольшие бобкэты и вилочные погрузчики, а при необходимости — фронтальные погрузчики и большие самосвалы.
- Затем мусор удаляется.
- Компания Wolf Paving перерабатывает старый асфальт и бетон на наших асфальтовых заводах, превращая разрушающиеся отходы в прочный, пригодный для использования новый асфальт. Фактически, компания Wolf Paving обычно перерабатывает почти 100% материалов, удаляемых с строительной площадки, что делает выбор в пользу использования Wolf Paving экологичным и безвредным для окружающей среды.
С чистого листа технология помогает специалистам по асфальту Wolf Paving подготовить поверхность для надлежащего отвода воды.
- Используя направляющие с лазерным наведением и автоматические автогрейдеры, команда Wolf Paving выравнивает поверхность, на которой будет производиться укладка, чтобы обеспечить надлежащий сток воды.
- Надлежащий дренаж воды жизненно важен для вашего асфальта, поскольку вода является основной причиной повреждений, включая выбоины, трещины и вздутие.
Вы никогда не догадаетесь, что самой важной частью вашего нового асфальтового покрытия на самом деле является основание.
- Основание обеспечивает устойчивую поверхность для поддержки нового дорожного покрытия.
- Подложка представляет собой барьер от мороза, помогающий уменьшить зимние повреждения из-за замерзания и оттаивания.
- Во время установки важными этапами являются толщина основания, устойчивость основания и уплотнение. Если основание не уплотнено должным образом, асфальтовое покрытие наверху не прослужит долгие годы.

Шаг 4. Контрольный валик, подрезка и ремонт основания
После того, как подложка полностью выровнена и уплотнена, компания Wolf Paving выполняет дополнительный этап, называемый контрольным валиком, чтобы обеспечить прочность основания и его готовность поддерживать новый асфальт.
- Контрольный валик Wolf Paving включает вождение четырехосного самосвала, загруженного 72 000 фунтов, ряд за рядом по всей поверхности.
- Если гравий изгибается более чем на дюйм под весом грузовика, это означает, что основание не поддерживается должным образом.
Если контрольный валик обнаружит мягкие участки основания, Wolf Paving произведет необходимый ремонт поврежденных участков, чтобы обеспечить поддержку всего основания.
- Подрезку можно использовать для ремонта мягких мест. Этот процесс включает в себя копание ниже поверхности на 2 или 3 фута и замену нижележащей мягкой глины или почвы более прочным заполнителем.
- Wolf Paving также предлагает альтернативные варианты, такие как использование геосетки вместо подрезки. Использование геосетки включает в себя копание всего на 16 дюймов ниже поверхности и укладку сетки для соединения основных материалов.Это создает прочную опорную структуру для нового асфальта за долю стоимости традиционной подрезки.
- Компания Wolf Paving также впервые применила новый процесс, называемый вспашкой. Этот процесс включает в себя подрезание основания, но вместо удаления всей мягкой глины и почвы его смешивают с добавленным заполнителем для повышения прочности скомпрометированных участков.
 Использование геосетки включает в себя копание всего на 16 дюймов ниже поверхности и укладку сетки для соединения основных материалов.Это создает прочную опорную структуру для нового асфальта за долю стоимости традиционной подрезки.
Использование геосетки включает в себя копание всего на 16 дюймов ниже поверхности и укладку сетки для соединения основных материалов.Это создает прочную опорную структуру для нового асфальта за долю стоимости традиционной подрезки.После укладки подложки и выявления и ремонта любых мягких участков настало время добавить вяжущее.
- Связующий слой представляет собой крупный заполнитель, смешанный с маслом, что делает его очень прочным и долговечным.
- Связующий слой можно рассматривать как прочность любого нового асфальтового покрытия.
После установки поддерживающих конструкций нового асфальтового покрытия добавляется верхний слой свежего асфальта, чтобы обеспечить чистоту и плавность движения.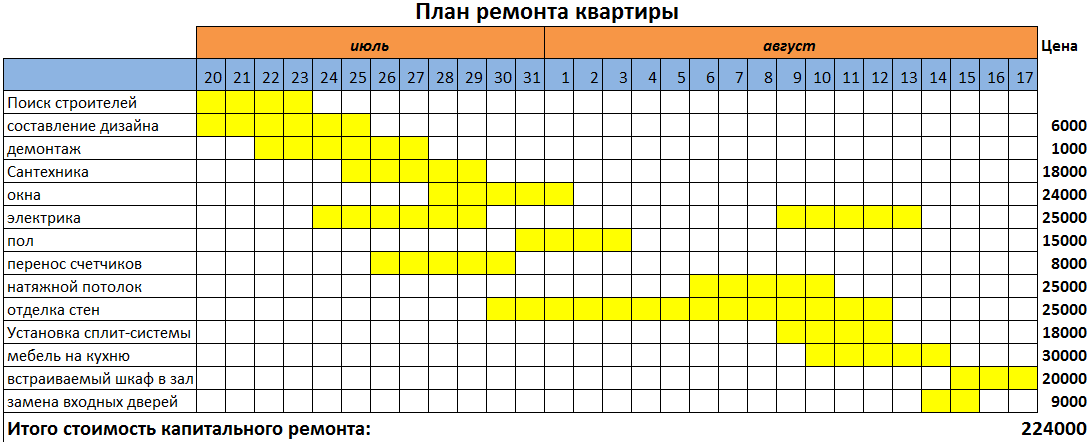
- Поверхностный асфальт состоит из мелкого заполнителя, песка и нефти.
- Эта комбинация материалов создает угольно-черный асфальт, который при правильной укладке обеспечивает плавность хода и блестящую, привлекательную поверхность.
Очень редко укладывают асфальтовое покрытие, которое не соединяется с существующими проездами, проезжей частью или парковками. Таким образом, подрядчики по укладке асфальта должны найти способ сгладить переход от старого покрытия к новому.
- Стыковые соединения представляют собой места, где старый асфальт или бетон встречаются с новым асфальтовым покрытием.
- Эти переходные участки требуют особого внимания, чтобы обеспечить надлежащий уклон и сток воды.
- Стыковые соединения важны для того, чтобы водители и пешеходы не замечали разницы в поверхностях.
После укладки асфальта и стыковых швов вся поверхность выравнивается и уплотняется.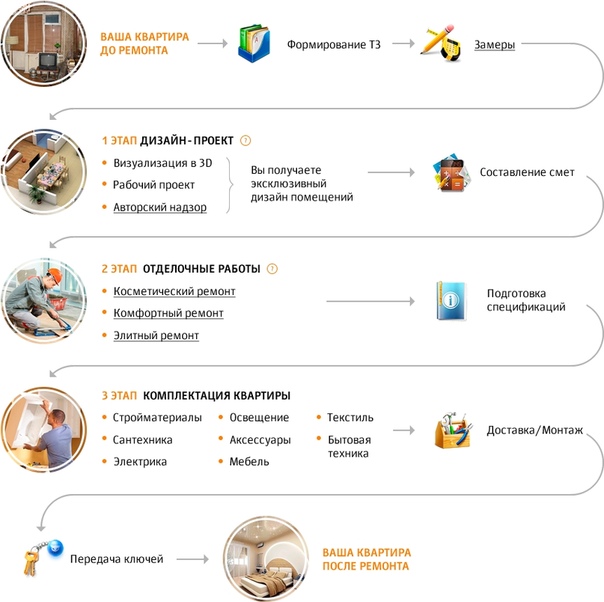
- С помощью роликовой тележки новое асфальтовое покрытие уплотняется и выравнивается.
- Этот этап процесса гарантирует, что на гладкой новой поверхности не останется мелких выпуклостей заполнителя или камня.
Ваш подрядчик по укладке асфальта должен выполнить следующие шаги по установке, чтобы убедиться, что ваше новое асфальтовое покрытие прослужит долгие годы и будет служить вашему дому, бизнесу, городу или поселку.
Но помните, что процесс установки должен следовать только после надлежащего осмотра поверхности и полной оценки. Взгляните на статьи ниже, которые полностью объясняют эти процессы.
Или нажмите здесь, чтобы перейти на нашу страницу ресурса «Все, что вам нужно знать о асфальтировании».
НАЖМИТЕ ЗДЕСЬ: Запросите бесплатную оценку сейчас для вашего следующего проекта по укладке тротуарной плитки
Благодаря 75-летнему опыту работы с асфальтом, от автомагистралей до подъездных дорог, мы являемся вашим местным поставщиком для всех ваших потребностей в асфальтировании.
Чтобы получить дополнительную информацию о Wolf Paving или запросить бесплатную оценку вашего следующего проекта по укладке тротуарной плитки, свяжитесь с одним из наших двух офисов. Если вы живете или работаете в районе Милуоки, позвоните нам по телефону 262-965-2121.В районе Мэдисона позвоните нам по телефону 608-249-7931.
Последовательность установки – обзор
11.1 Общие положения
На внешнем континентальном шельфе США правила требуют, чтобы арендатор демонтировал все конструкции и связанные с ними подводные препятствия в течение одного года после прекращения аренды (Федеральный реестр, 2002 г.). Морские операции по выводу из эксплуатации более сложны и значительно дороже, чем работы на суше, из-за логистических проблем, связанных с работой в изолированных средах в водах различной глубины и погодных условий.По мере увеличения глубины воды размер конструкции увеличивается, и для подъемных операций требуются специализированные морские суда, проекты удаляются от берега и требуют больше времени на планирование и выполнение, а для выполнения работ доступно меньше судов, что увеличивает стоимость проекта и неопределенность.
Операции по выводу из эксплуатации выполняются, по существу, в обратной последовательности последовательности установки, но общая продолжительность работ обычно намного короче, а основные этапы проекта менее ограничены (Ayoade, 2002).Выбор оборудования и процедур при выводе из эксплуатации зависит от многих из тех же факторов, которые влияют на выбор для установки, включая доступность и стоимость оборудования, а также характер и последовательность операций.
После установки экспортного трубопровода скорость потока близка к максимальной, а загрузка линии высока. Через некоторое время, когда добыча на месторождении снизится, скорость потока и использование трубопровода также снизятся. Если в регионе будут обнаружены новые месторождения и экономически выгодна их привязка к платформе, и будет достигнуто соглашение с владельцем сооружения (если он отличается от владельца месторождения), экспортная линия будет продолжать использоваться.Однако в какой-то момент вблизи платформы не будет новых открытий, или расстояние до платформы будет слишком велико для обратной связи, и коэффициенты нагрузки на линии снизятся.
Существует несколько признаков того, что срок службы газопровода приближается к концу (вставка 11.1). Если у трубопровода низкий доход или он отсутствует, нет твердых контрактов, нет клиентов или есть только небольшое количество клиентов с перебоями в обслуживании, дорогостоящий ремонт, проблемы со очисткой скребками, коррозия и эксплуатационные проблемы, то, скорее всего, срок службы трубопровода подходит к концу. жизнь.
Блок 11.1Вопросы жизненного цикла и индикаторы окончания срока службы
Эксплуатационные соображения
Коэффициент нагрузки определяется как объем пропускной способности, деленный на пропускную способность трубопровода за определенный период времени. В течение первых нескольких лет после установки коэффициенты нагрузки обычно превышают 90 %, поскольку экспортные трубопроводы спроектированы так, чтобы соответствовать максимальному ожидаемому расходу воды из сооружения. Коэффициент загрузки 10% или менее обычно считается низким и, хотя и не свидетельствует о скором выводе из эксплуатации, обычно является индикатором будущей деятельности.
При низких коэффициентах загрузки фиксированные эксплуатационные расходы распределяются на уменьшающиеся объемы, что приводит к тому, что оператор трубопровода потенциально стремится повысить ставки для существующих клиентов. Если трубопровод регулируется FERC, предлагаемое повышение тарифной ставки требует одобрения FERC. Если трубопровод не регулируется FERC, у грузоотправителя не остается иного выхода, кроме как платить более высокие ставки. Хотя FERC не обладает юрисдикцией для обеспечения разумности ставок на нефтепроводах, трубопроводы OCS должны по-прежнему придерживаться политики недискриминации и открытого доступа (см. Приложение C).Раздел 5(f) Закона о землях внешнего континентального шельфа 1953 года требует, чтобы все трубопроводы OCS «обеспечивали открытый и недискриминационный доступ как для владельцев, так и для невладельцев».
Вопросы владения
Если экспортную линию эксплуатируют владельцы скважин или дочерняя или аффилированная компания владельцев скважин, линия, скорее всего, останется в эксплуатации до тех пор, пока она не перестанет быть коммерческой, но если третья сторона будет эксплуатировать экспортную линию, выгода от обслуживания линия ближе к концу срока службы месторождения будет значительно отличаться от владельцев ресурсов, потому что доходы трубопроводной компании полностью основаны на спросе (дебитах) и товарных сборах за использование трубопровода. Когда дебиты снижаются, обычно следуют доходы, и владельцы трубопроводов не получают никаких преимуществ от владения ресурсами.
Когда дебиты снижаются, обычно следуют доходы, и владельцы трубопроводов не получают никаких преимуществ от владения ресурсами.
Проблема с потоками отличается, потому что владельцы ресурсов почти всегда владеют потоками и управляют ими. По мере снижения добычи на месторождении использование выкидных трубопроводов, которые доставляют продукцию скважины на предприятие для переработки, сокращается и, в конечном итоге, станет нерентабельным в эксплуатации, но поскольку большинство выкидных трубопроводов принадлежит и эксплуатируется владельцем (владельцами) подводных скважин, конфликт не возникает. представляет интерес.Если скважина работает и система экономична в эксплуатации, выкидная линия будет обслуживаться для поддержания добычи.
Для линии, управляемой третьей стороной, могут рассматриваться дела о ставках, и повышенные ставки передаются клиентам для поддержания работы, но эти разбирательства требуют времени, а результаты неясны. Владелец скважины, транспортирующей газ по трубопроводу FERC, может возражать против отказа от линии, потому что это может привести к выбросу углеводородов и ограничению вариантов будущей разработки, и, конечно же, владельцы скважин предпочли бы, чтобы линия оставалась в эксплуатации как можно дольше, независимо от расходы оператора трубопровода. FERC пытается сбалансировать эти конкурирующие интересы, используя руководящие принципы, называемые общественным удобством и необходимостью.
FERC пытается сбалансировать эти конкурирующие интересы, используя руководящие принципы, называемые общественным удобством и необходимостью.
Индикаторы окончания срока службы
Трубопроводы и стояки проверяются по графику, установленному правилами BSEE, обычно раз в 3-5 лет. Если трубы теряют более половины своей толщины, их необходимо заменить или отключить. Когда скорость потока низкая, очистка скребками становится более сложной, поскольку скребкам требуется давление, чтобы протолкнуть их через трубопровод, а покупка газа может быть дорогой, или подача газа может быть недоступна.Уменьшение скребков ускоряет коррозию и приводит к проблемам безопасности и экологии, в зависимости от возраста и состояния линии.
Большинство трубопроводов в Мексиканском заливе оставлены на месте, а выведенные из эксплуатации участки трубопроводов не удаляются с морского дна. Удаляются только небольшие участки трубопровода, как правило, стояки и повороты труб, или, если трубопровод поврежден или протекает, поврежденный участок необходимо отремонтировать или заменить перед выводом из эксплуатации. Правила требуют, чтобы концы трубопровода были закопаны на 3 фута ниже морского дна или покрыты бетонным матом.Процедуры на глубоководье аналогичны процедурам на мелководье, но более сложны (Burke and Stokes, 2015; Phillip et al., 2014). Процедуры вывода из эксплуатации в Мексиканском заливе также аналогичны, но не идентичны процедурам в других прибрежных регионах, таких как Северное море (DECC, 2011), где различия в режимах регулирования и морском дне создают различия в практике вывода из эксплуатации.
Правила требуют, чтобы концы трубопровода были закопаны на 3 фута ниже морского дна или покрыты бетонным матом.Процедуры на глубоководье аналогичны процедурам на мелководье, но более сложны (Burke and Stokes, 2015; Phillip et al., 2014). Процедуры вывода из эксплуатации в Мексиканском заливе также аналогичны, но не идентичны процедурам в других прибрежных регионах, таких как Северное море (DECC, 2011), где различия в режимах регулирования и морском дне создают различия в практике вывода из эксплуатации.
В районах Внешнего континентального шельфа, в которых определены значительные запасы наносов, Бюро по управлению энергетикой океана требует, чтобы трубопроводы выводились из эксплуатации путем удаления, что является более длительной и сложной операцией.Компании должны получить разрешение Бюро по безопасности и охране окружающей среды перед операцией по отказу от полосы отчуждения, а поскольку во время операций по выводу из эксплуатации взрывные работы не проводятся, обычно не требуется дополнительных экологических разрешений.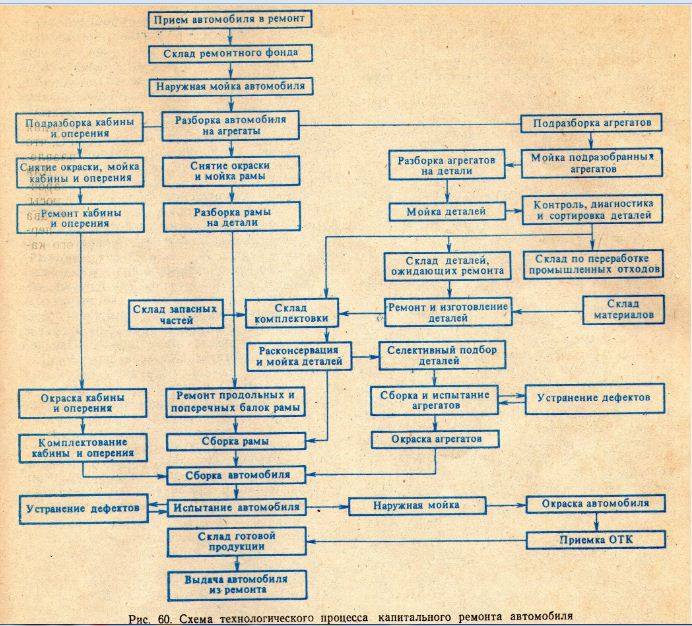 Закон об управлении прибрежной зоной в Луизиане требует пересмотра всех операций по выводу трубопроводов из эксплуатации и согласования с государством в каждом конкретном случае.
Закон об управлении прибрежной зоной в Луизиане требует пересмотра всех операций по выводу трубопроводов из эксплуатации и согласования с государством в каждом конкретном случае.
Другие штаты Мексиканского залива различаются по своей прибрежной зоне и политике управления трубопроводами.Конечные точки трубопровода, которые полностью находятся в пределах федеральных вод, подлежат категорическому исключению из требований по подготовке экологической оценки или отчета о воздействии на окружающую среду, но трубопроводы, пересекающие воды штата и федеральные воды, требуют дополнительной подготовки и проверки, обычно включающей отчет о воздействии на окружающую среду. Трубопроводы, которые не подпадают под действие правил FERC, требуют только разрешения BSEE и разрешения на отказ.
Трубопроводы, по которым проходят двухфазные жидкости (нефть-газ, газ-конденсат), более подвержены коррозии, чем те, по которым проходят однофазные жидкости.Сырая нефть с высоким содержанием водяного пара и серы, а также газ с высоким содержанием серы, CO 2 и водяным паром, являются коррозионно-активными, и чем ниже давление потока, тем более коррозионным является воздействие. Удельное сопротивление морского дна, соленость воды и состав морского дна могут способствовать коррозионной активности и влиять на вероятность активной коррозии.
Удельное сопротивление морского дна, соленость воды и состав морского дна могут способствовать коррозионной активности и влиять на вероятность активной коррозии.
Что такое редактирование генома и CRISPR-Cas9?: MedlinePlus Genetics
Редактирование генома (также называемое редактированием генов) — это группа технологий, которые дают ученым возможность изменять ДНК организма.Эти технологии позволяют добавлять, удалять или изменять генетический материал в определенных местах генома. Было разработано несколько подходов к редактированию генома. Недавняя система известна как CRISPR-Cas9, что является сокращением от сгруппированных регулярно расположенных коротких палиндромных повторов и CRISPR-ассоциированного белка 9. Система CRISPR-Cas9 вызвала большой интерес в научном сообществе, потому что она быстрее, дешевле, больше. точным и более эффективным, чем другие существующие методы редактирования генома.
CRISPR-Cas9 был адаптирован из естественной системы редактирования генома бактерий.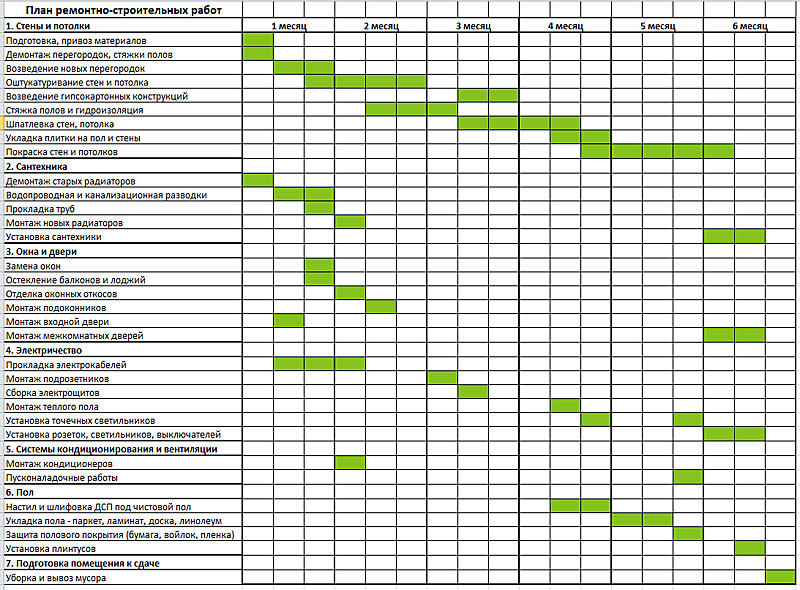 Бактерии захватывают фрагменты ДНК вторгшихся вирусов и используют их для создания сегментов ДНК, известных как массивы CRISPR. Массивы CRISPR позволяют бактериям «запоминать» вирусы (или близкородственные). Если вирусы атакуют снова, бактерии производят сегменты РНК из массивов CRISPR для нацеливания на ДНК вирусов. Затем бактерии используют Cas9 или аналогичный фермент, чтобы разрезать ДНК на части, что отключает вирус.
Бактерии захватывают фрагменты ДНК вторгшихся вирусов и используют их для создания сегментов ДНК, известных как массивы CRISPR. Массивы CRISPR позволяют бактериям «запоминать» вирусы (или близкородственные). Если вирусы атакуют снова, бактерии производят сегменты РНК из массивов CRISPR для нацеливания на ДНК вирусов. Затем бактерии используют Cas9 или аналогичный фермент, чтобы разрезать ДНК на части, что отключает вирус.
Аналогично работает система CRISPR-Cas9 в лаборатории. Исследователи создают небольшой фрагмент РНК с короткой «направляющей» последовательностью, которая прикрепляется (связывается) к определенной целевой последовательности ДНК в геноме. РНК также связывается с ферментом Cas9. Как и у бактерий, модифицированная РНК используется для распознавания последовательности ДНК, а фермент Cas9 разрезает ДНК в нужном месте. Хотя чаще всего используется фермент Cas9, можно использовать и другие ферменты (например, Cpf1). После разрезания ДНК исследователи используют собственный механизм репарации ДНК клетки, чтобы добавлять или удалять фрагменты генетического материала или вносить изменения в ДНК, заменяя существующий сегмент индивидуальной последовательностью ДНК.
Редактирование генома представляет большой интерес для профилактики и лечения заболеваний человека. В настоящее время большинство исследований по редактированию генома проводится для понимания болезней с использованием клеток и животных моделей. Ученые все еще работают над тем, чтобы определить, является ли этот подход безопасным и эффективным для использования на людях. Он изучается в исследованиях широкого спектра заболеваний, включая моногенные расстройства, такие как муковисцидоз, гемофилия и серповидно-клеточная анемия. Он также обещает лечение и профилактику более сложных заболеваний, таких как рак, болезни сердца, психические заболевания и инфекция, вызванная вирусом иммунодефицита человека (ВИЧ).
Этические проблемы возникают, когда редактирование генома с использованием таких технологий, как CRISPR-Cas9, используется для изменения геномов человека. Большинство изменений, вносимых при редактировании генома, ограничиваются соматическими клетками, то есть клетками, отличными от яйцеклеток и сперматозоидов. Эти изменения затрагивают только определенные ткани и не передаются из поколения в поколение. Однако изменения, внесенные в гены яйцеклеток или сперматозоидов (клетки зародышевой линии) или в гены эмбриона, могут быть переданы будущим поколениям. Редактирование генома зародышевых клеток и эмбрионов поднимает ряд этических проблем, в том числе вопрос о том, допустимо ли использовать эту технологию для улучшения нормальных человеческих черт (таких как рост или интеллект).Из соображений этики и безопасности редактирование генома зародышевых клеток и эмбрионов в настоящее время является незаконным во многих странах.
Эти изменения затрагивают только определенные ткани и не передаются из поколения в поколение. Однако изменения, внесенные в гены яйцеклеток или сперматозоидов (клетки зародышевой линии) или в гены эмбриона, могут быть переданы будущим поколениям. Редактирование генома зародышевых клеток и эмбрионов поднимает ряд этических проблем, в том числе вопрос о том, допустимо ли использовать эту технологию для улучшения нормальных человеческих черт (таких как рост или интеллект).Из соображений этики и безопасности редактирование генома зародышевых клеток и эмбрионов в настоящее время является незаконным во многих странах.
Статьи из научных журналов для дополнительного чтения
Ормонд К.Е.(1), Мортлок Д.П.(2), Скоулз Д.Т.(3), Бомбард Y(4), Броди Л.К.(5), Фосетт В.А.(6), Гарнизон Н.А.(7), Херчер Л.(8), Исаси Р (9), Миддлтон А (10), Мусунуру К (11), Шрайнер Д (12), Вирани А (13), Янг КЭ (3). Редактирование генома зародышевой линии человека. Am J Hum Genet. 2017 3 августа; 101 (2): 167–176. PubMed: 28777929. Бесплатный полный текст доступен в PubMed Central: PMC5544380.
PubMed: 28777929. Бесплатный полный текст доступен в PubMed Central: PMC5544380.
Гупта Р.М., Мусунуру К. Расширение набора инструментов генетического редактирования: ZFN, TALEN и CRISPR-Cas9. Джей Клин Инвест. 2014 Октябрь; 124 (10): 4154-61. DOI: 10.1172/JCI72992. Epub 2014 1 октября. Обзор. PubMed: 25271723. Бесплатный полный текст доступен в PubMed Central: PMC41.
Хсу П.Д., Ландер Э.С., Чжан Ф. Разработка и применение CRISPR-Cas9 для инженерии генома. Клетка. 2014 5 июня; 157 (6): 1262-78. doi:10.1016/j.cell.2014.05.010. Обзор. PubMed: 246. Бесплатный полный текст доступен в PubMed Central: PMC4343198.
Комор А.С., Бадран А.Х., Лю Д.Р. Технологии на основе CRISPR для манипулирования геномами эукариот. Клетка. 20 апр. 2017; 169(3):559. doi:10.1016/j.cell.2017.04.005. ПабМед: 28431253.
Ландер ЕС. Герои CRISPR. Клетка. 2016 14 января; 164 (1-2): 18-28. doi:10.1016/j.cell.2015.12.041. Обзор. PubMed: 26771483.
Редактирование генов при кистозном фиброзе
Как работает редактирование генов?
Ген – это серия букв ДНК, которая содержит инструкции по созданию белка. Поскольку ДНК имеет решающее значение для клетки, у клетки есть механизмы для исправления повреждений ДНК. Редактирование генов использует собственный механизм восстановления ДНК клетки, чтобы исправить мутацию в ДНК клетки. В отличие от генозаместительной терапии, редактирование генов исправляет мутации в собственной ДНК человека.
Люди с муковисцидозом имеют мутацию в обеих копиях гена регулятора проводимости мембраны муковисцидоза (CFTR). Чтобы исправить мутации CFTR в чьей-то ДНК, инструменты, необходимые для редактирования генов, должны проникнуть внутрь клеток человека, что очень сложно.
После того, как инструмент попал в ядро клетки, он должен быть в состоянии найти серию из примерно 20 букв ДНК, которые отмечают место мутации CFTR из трех миллиардов букв в геноме человека. Затем ДНК нужно разорвать прямо рядом с мутацией в гене CFTR.
Посмотрите это видео, чтобы увидеть, как этот процесс может работать.
Типы инструментов редактирования генов
Ученые обнаружили несколько различных инструментов, которые могут найти определенную серию букв в геноме и разрушить ДНК в этом месте. Самый универсальный инструмент для редактирования генов называется CRISPR. CRISPR часто предпочтительнее, потому что он недорог по сравнению с другими методами и его легче всего настроить, то есть легко указать, какие серии букв ДНК CRISPR будет искать в геноме.
Самый универсальный инструмент для редактирования генов называется CRISPR. CRISPR часто предпочтительнее, потому что он недорог по сравнению с другими методами и его легче всего настроить, то есть легко указать, какие серии букв ДНК CRISPR будет искать в геноме.
Инструменты редактирования гена CRISPR включают в себя «направляющую», которая находит мутантную последовательность в гене CFTR, матрицу с правильным сегментом букв ДНК и «ножницы», которые разрывают ДНК пациента в месте мутации.
Как только инструменты проникают в клетку и достигают мутировавшей последовательности ДНК, ножницы вырезают мутацию. Это повреждение привлекает внимание механизма репарации ДНК клетки, который затем использует матрицу, чтобы исправить разрыв в ДНК. Это навсегда исправляет мутацию в этой клетке. Этот процесс редактирования генов может исправлять одну мутацию за раз или группы подобных мутаций, в зависимости от того, как мутации расположены в ДНК.
Посмотрите это видео, чтобы узнать, как работает редактирование CRISPR/Cas9.
Существуют и другие инструменты, которые можно использовать для редактирования генов, включая белки, называемые TALEN, мегануклеазы и нуклеазы цинковых пальцев. Все они работают аналогично CRISPR, но менее популярны, потому что их сложнее настроить и требуют от исследователей гораздо больше времени и опыта.
Проблемы редактирования генов для CF
Основным преимуществом редактирования генов является то, что изменения, которые оно вносит в ДНК, являются постоянными. Людям может потребоваться только одно лечение с помощью инструмента для редактирования генов, чтобы оказать длительное влияние на их болезнь.Тем не менее, еще предстоит решить множество проблем, прежде чем редактирование генов можно будет использовать для лечения муковисцидоза.
Например, инструменты редактирования генов можно настроить для поиска конкретной мутации CFTR, но существует множество мутаций, вызывающих муковисцидоз. Исследователи изучают эффективность и действенность различных методов в лаборатории, которые позволили бы процессу коррекции исправлять где угодно, от одной мутации до всех мутаций.
Редактирование вне сайта также представляет риск. Теоретически редактирование генов должно быть очень точной терапией, а это означает, что инструмент редактирования генов должен разрушать ДНК человека только в том месте, которое он должен был найти.Для муковисцидоза инструмент будет разработан для поиска места мутации CFTR.
На практике редактирование генов несовершенно. Иногда инструменты редактирования генов разрывают ДНК в неправильном месте генома. Такая ошибка может привести к новым мутациям в других генах и вызвать непредвиденные последствия, такие как повышенный риск развития рака. По этой причине каждый инструмент редактирования генов должен оцениваться индивидуально, чтобы определить, достаточно ли он точен для использования у пациентов.
Современные технологии редактирования генов полагаются на собственный механизм восстановления ДНК клетки, чтобы исправить разрыв в ДНК, вызванный инструментом редактирования генов.Однако механизм репарации ДНК клетки может завершить свою работу только в том случае, если клетка претерпевает цикл клеточного деления (одна клетка делится на две новые клетки). У взрослых небольшая популяция стволовых клеток дыхательных путей регулярно подвергается клеточному делению; однако большинство клеток в таких органах, как легкие, не делятся. Это может означать, что редактирование генов не сможет исправить мутации CFTR во многих клетках легких, если только стволовые клетки не будут специально нацелены. Разрабатываются новые технологии, которые могут работать даже в неделящихся клетках.Эти новые технологии могут позволить редактированию генов работать в большем количестве клеток в легких.
У взрослых небольшая популяция стволовых клеток дыхательных путей регулярно подвергается клеточному делению; однако большинство клеток в таких органах, как легкие, не делятся. Это может означать, что редактирование генов не сможет исправить мутации CFTR во многих клетках легких, если только стволовые клетки не будут специально нацелены. Разрабатываются новые технологии, которые могут работать даже в неделящихся клетках.Эти новые технологии могут позволить редактированию генов работать в большем количестве клеток в легких.
Редактирование генов является областью очень активных исследований и приближается к клиническим испытаниям для лечения нескольких заболеваний крови, таких как серповидно-клеточная анемия. Эти заболевания являются хорошими мишенями для редактирования генов, потому что клетки крови можно извлечь из организма и лечить с помощью инструментов редактирования генов в лаборатории. Как только инструменты редактирования генов исправят мутации, клетки крови можно будет вернуть в организм.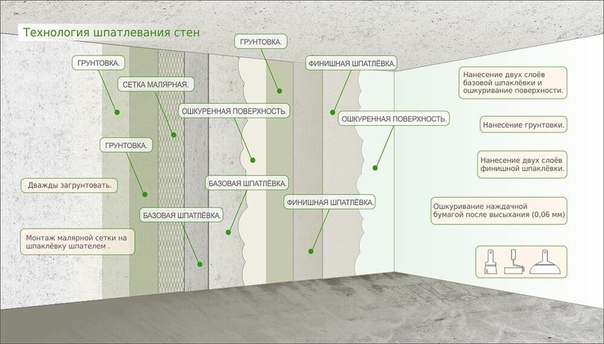
Лечение такого заболевания, как муковисцидоз, которое поражает легкие и другие внутренние органы, намного сложнее, потому что очень сложно ввести инструменты редактирования генов в клетки легких, процесс, называемый доставкой генов. Редактирование генов для CF в настоящее время тестируется на клетках и животных, и пройдет несколько лет, прежде чем его можно будет безопасно протестировать на людях.
Мутации, репарация и рекомбинация — геномы
Геномы представляют собой динамические объекты, которые изменяются с течением времени в результате кумулятивных эффектов небольших изменений последовательности, вызванных мутациями, и более масштабных перестроек, возникающих в результате рекомбинации.Мутация и рекомбинация могут быть определены как процессы, которые приводят к изменениям в геноме , но они не связаны между собой, и мы должны проводить между ними четкое различие:
Как мутация, так и рекомбинация могут иметь серьезные последствия для клетки, в которой они происходят.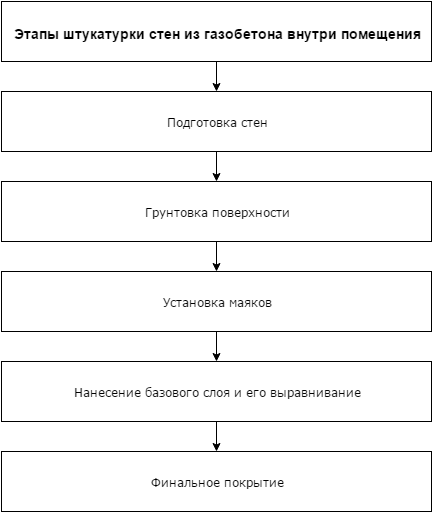 Мутация в ключевом гене может привести к гибели клетки, если белок, кодируемый мутантным геном, дефектен (раздел 14.1.2), а некоторые события рекомбинации приводят к определяющим изменениям в биохимических возможностях клетки, например, путем определения тип спаривания дрожжевой клетки или иммунологические свойства В- или Т-лимфоцитов млекопитающих.Другие события мутации и рекомбинации оказывают менее значительное влияние на фенотип клетки, а многие вообще не влияют. Как мы увидим в главе 15, все нелетальные события потенциально могут способствовать эволюции генома, но для этого они должны быть унаследованы при воспроизведении организма. В одноклеточном организме, таком как бактерия или дрожжи, все изменения генома, которые не являются летальными или обратимыми, наследуются дочерними клетками и становятся постоянными чертами линии, происходящей от исходной клетки, в которой произошло изменение.В многоклеточном организме только те события, которые происходят в зародышевых клетках, имеют отношение к эволюции генома.
Мутация в ключевом гене может привести к гибели клетки, если белок, кодируемый мутантным геном, дефектен (раздел 14.1.2), а некоторые события рекомбинации приводят к определяющим изменениям в биохимических возможностях клетки, например, путем определения тип спаривания дрожжевой клетки или иммунологические свойства В- или Т-лимфоцитов млекопитающих.Другие события мутации и рекомбинации оказывают менее значительное влияние на фенотип клетки, а многие вообще не влияют. Как мы увидим в главе 15, все нелетальные события потенциально могут способствовать эволюции генома, но для этого они должны быть унаследованы при воспроизведении организма. В одноклеточном организме, таком как бактерия или дрожжи, все изменения генома, которые не являются летальными или обратимыми, наследуются дочерними клетками и становятся постоянными чертами линии, происходящей от исходной клетки, в которой произошло изменение.В многоклеточном организме только те события, которые происходят в зародышевых клетках, имеют отношение к эволюции генома. Изменения в геномах соматических клеток не важны в эволюционном смысле, но они будут иметь биологическую значимость, если приведут к вредному фенотипу, влияющему на здоровье организма.
Изменения в геномах соматических клеток не важны в эволюционном смысле, но они будут иметь биологическую значимость, если приведут к вредному фенотипу, влияющему на здоровье организма.
Вставка 14.1
Терминология для описания точечных мутаций. Точечные мутации также называют простыми мутациями или мутациями с одним сайтом. Иногда их называют мутациями замещения, но это может привести к путанице, потому что для генетика-эволюциониста «замещение» (подробнее…)
14.1. Мутации
Что касается мутаций, мы должны рассмотреть следующие вопросы: как они возникают; влияние, которое они оказывают на геном и на организм, в котором находится геном; может ли клетка увеличивать частоту мутаций и индуцировать запрограммированные мутации при определенных обстоятельствах; и как мутации исправляются.
14.1.1. Причины мутаций
Мутации возникают двумя путями:
- Некоторые мутации представляют собой спонтанных ошибок в репликации, которые ускользают от корректирующей функции ДНК-полимераз, синтезирующих новые полинуклеотиды на репликационной вилке (Раздел 13. 2.2). Эти мутации называются несоответствиями , потому что они представляют собой положения, в которых нуклеотид, вставленный в дочерний полинуклеотид, не совпадает, путем спаривания оснований, с нуклеотидом в соответствующем положении в матричной ДНК (1). Если несоответствие сохраняется в дочерней двойной спирали, то 90 431 одна 90 432 внучатых молекул, образующихся в ходе следующего раунда репликации ДНК, будут нести постоянную двухцепочечную версию мутации.
- Другие мутации возникают из-за того, что мутаген вступает в реакцию с исходной ДНК, вызывая структурное изменение, влияющее на способность измененного нуклеотида образовывать пары оснований.Обычно это изменение затрагивает только одну нить родительской двойной спирали, поэтому только одна из дочерних молекул несет мутацию, но две из внучатых молекул, образующихся в ходе следующего раунда репликации, будут иметь ее (2).
 2.2). Эти мутации называются несоответствиями , потому что они представляют собой положения, в которых нуклеотид, вставленный в дочерний полинуклеотид, не совпадает, путем спаривания оснований, с нуклеотидом в соответствующем положении в матричной ДНК (1). Если несоответствие сохраняется в дочерней двойной спирали, то 90 431 одна 90 432 внучатых молекул, образующихся в ходе следующего раунда репликации ДНК, будут нести постоянную двухцепочечную версию мутации.
2.2). Эти мутации называются несоответствиями , потому что они представляют собой положения, в которых нуклеотид, вставленный в дочерний полинуклеотид, не совпадает, путем спаривания оснований, с нуклеотидом в соответствующем положении в матричной ДНК (1). Если несоответствие сохраняется в дочерней двойной спирали, то 90 431 одна 90 432 внучатых молекул, образующихся в ходе следующего раунда репликации ДНК, будут нести постоянную двухцепочечную версию мутации.Рисунок 14.2
Примеры мутаций. (A) Ошибка в репликации приводит к несоответствию в одной из дочерних двойных спиралей, в данном случае к изменению T-C, потому что один из As в матричной ДНК был неправильно скопирован. Когда несовпадающая молекула сама реплицируется, она дает (больше…)
Когда несовпадающая молекула сама реплицируется, она дает (больше…)
Ошибки репликации являются источником точечных мутаций
Если рассматривать исключительно химическую реакцию, комплементарное спаривание оснований не является особенно точным. Никто еще не придумал способ проведения зависимого от матрицы синтеза ДНК без помощи ферментов, но если бы этот процесс можно было проводить просто как химическую реакцию в пробирке, то полученный полинуклеотид, вероятно, имел бы точечные мутации в 5-м положении. –10 позиций из каждой сотни. Это соответствует частоте ошибок 5–10%, что было бы совершенно неприемлемо при репликации генома.Таким образом, зависимые от матрицы ДНК-полимеразы, осуществляющие репликацию ДНК, должны повысить точность процесса на несколько порядков. Это улучшение достигается двумя способами:
- ДНК-полимераза управляет процессом отбора нуклеотидов, который резко повышает точность зависимого от матрицы синтеза ДНК (). Этот процесс отбора, вероятно, действует на трех различных стадиях реакции полимеризации: дискриминация неправильного нуклеотида происходит, когда нуклеотид впервые связывается с ДНК-полимеразой, когда он перемещается в активный центр фермента и когда он присоединяется к ДНК-полимеразе. 3′-конец синтезируемого полинуклеотида.
- Точность синтеза ДНК повышается еще больше, если ДНК-полимераза обладает 3’→5′-экзонуклеазной активностью и, таким образом, способна удалять неправильный нуклеотид, который уклоняется от процесса отбора оснований и прикрепляется к 3′-концу нового полинуклеотид (см. ). Это называется вычиткой (раздел 13.2.2), но это название является неправильным, поскольку этот процесс не является активным механизмом проверки. Напротив, каждый этап синтеза полинуклеотида следует рассматривать как соревнование между полимеразной и экзонуклеазной функциями фермента, при этом полимераза обычно побеждает, поскольку она более активна, чем экзонуклеаза, по крайней мере, когда 3′-концевой нуклеотид является основанием. — в паре с шаблоном.Но активность полимеразы менее эффективна, если концевой нуклеотид не имеет пар оснований, что приводит к паузе в полимеризации, позволяющей преобладать экзонуклеазной активности, так что неправильный нуклеотид удаляется (см. ).
 3′-конец синтезируемого полинуклеотида.
3′-конец синтезируемого полинуклеотида. ).
).Рисунок 14.3
Механизмы обеспечения точности репликации ДНК. (A) ДНК-полимераза активно выбирает правильный нуклеотид для вставки в каждом положении. (B) Те ошибки, которые возникают, могут быть исправлены «вычиткой», если полимераза имеет (подробнее…)
Escherichia coli способна синтезировать ДНК с частотой ошибок всего 1 на 10 7 добавлений нуклеотидов. Интересно, что эти ошибки неравномерно распределены между двумя дочерними молекулами, продукт репликации отстающей цепи подвержен примерно в 20 раз большему количеству ошибок, чем репликация лидирующей цепи. Эта асимметрия может указывать на то, что ДНК-полимераза I, участвующая только в репликации отстающих цепей (раздел 13.2.2), обладает менее эффективной способностью к выбору оснований и корректуре по сравнению с ДНК-полимеразой III, основным репликационным ферментом (Francino and Ochman, 1997). ).
Не все ошибки, возникающие при синтезе ДНК, можно списать на ферменты-полимеразы: иногда ошибка возникает, даже если фермент добавляет «правильный» нуклеотид, тот, который образует пару оснований с матрицей. Это связано с тем, что каждое нуклеотидное основание может встречаться в виде любого из двух альтернативных таутомеров, структурных изомеров, которые находятся в динамическом равновесии. Например, тимин существует в виде двух таутомеров, кето и енола , причем отдельные молекулы иногда претерпевают переход от одного таутомера к другому.Равновесие сильно смещено в сторону формы кето , но время от времени енольная версия тимина встречается в матричной ДНК в точное время, когда проходит репликационная вилка. Это приведет к «ошибке», потому что енол -тимин образуют пары оснований с G, а не с A (). Та же проблема может возникнуть с аденином, редкий имино таутомер этого основания преимущественно образует пару с С, а с гуанином енол -гуанин образует пару с тимином.После репликации редкий таутомер неизбежно вернется к своей более распространенной форме, что приведет к несоответствию в дочерней двойной спирали.
Это связано с тем, что каждое нуклеотидное основание может встречаться в виде любого из двух альтернативных таутомеров, структурных изомеров, которые находятся в динамическом равновесии. Например, тимин существует в виде двух таутомеров, кето и енола , причем отдельные молекулы иногда претерпевают переход от одного таутомера к другому.Равновесие сильно смещено в сторону формы кето , но время от времени енольная версия тимина встречается в матричной ДНК в точное время, когда проходит репликационная вилка. Это приведет к «ошибке», потому что енол -тимин образуют пары оснований с G, а не с A (). Та же проблема может возникнуть с аденином, редкий имино таутомер этого основания преимущественно образует пару с С, а с гуанином енол -гуанин образует пару с тимином.После репликации редкий таутомер неизбежно вернется к своей более распространенной форме, что приведет к несоответствию в дочерней двойной спирали.
Рисунок 14.4
Влияние таутомерии на спаривание оснований. В каждом из этих трех примеров две таутомерные формы основания имеют разные свойства спаривания. Цитозин также имеет таутомеры амино и имино , но оба они связаны с гуанином.
В каждом из этих трех примеров две таутомерные формы основания имеют разные свойства спаривания. Цитозин также имеет таутомеры амино и имино , но оба они связаны с гуанином.
Как указано выше, частота ошибок при синтезе ДНК в E.coli составляет 1 из 10 7 . Общая частота ошибок для репликации генома E. coli составляет всего от 1 на 10 10 до 1 на 10 11 , улучшение по сравнению с частотой ошибок полимеразы является результатом системы репарации несоответствия (раздел 14.2. 3), который сканирует только что реплицированную ДНК в поисках положений, в которых основания не спарены, и, следовательно, исправляет несколько ошибок, которые делают репликационные ферменты. Подразумевается, что только одна неисправленная ошибка репликации возникает каждые 1000 раз, когда ошибка E.coli копируется геном.
Ошибки репликации также могут привести к мутациям вставки и удаления
Не все ошибки репликации являются точечными мутациями. Аберрантная репликация также может привести к вставке небольшого количества дополнительных нуклеотидов в синтезируемый полинуклеотид или к тому, что некоторые нуклеотиды в матрице не будут скопированы. Инсерции и делеции часто называют мутациями со сдвигом рамки считывания , потому что, когда они происходят в кодирующей области, они могут привести к сдвигу в рамке считывания, используемой для трансляции белка, указанного геном (см. Ресурсы).Однако неправильно использовать «сдвиг рамки» для описания всех вставок и делеций, поскольку они могут встречаться где угодно, а не только в генах, и не все вставки или делеции в кодирующих областях приводят к сдвигу рамки считывания: вставка или удаление трех нуклеотидов или множественные из трех, просто добавляет или удаляет кодоны или части соседних кодонов, не затрагивая рамку считывания.
Аберрантная репликация также может привести к вставке небольшого количества дополнительных нуклеотидов в синтезируемый полинуклеотид или к тому, что некоторые нуклеотиды в матрице не будут скопированы. Инсерции и делеции часто называют мутациями со сдвигом рамки считывания , потому что, когда они происходят в кодирующей области, они могут привести к сдвигу в рамке считывания, используемой для трансляции белка, указанного геном (см. Ресурсы).Однако неправильно использовать «сдвиг рамки» для описания всех вставок и делеций, поскольку они могут встречаться где угодно, а не только в генах, и не все вставки или делеции в кодирующих областях приводят к сдвигу рамки считывания: вставка или удаление трех нуклеотидов или множественные из трех, просто добавляет или удаляет кодоны или части соседних кодонов, не затрагивая рамку считывания.
Рисунок 14.12
Делеционные мутации. В верхней последовательности удалены три нуклеотида, составляющие один кодон.Это укорачивает полученный белковый продукт на одну аминокислоту, но не влияет на остальную часть его последовательности. В нижней части удален один нуклеотид. (подробнее…)
В нижней части удален один нуклеотид. (подробнее…)
Инсерционные и делеционные мутации могут затрагивать все части генома, но особенно распространены, когда матричная ДНК содержит короткие повторяющиеся последовательности, такие как обнаруженные в микросателлитах (раздел 2.4.1). Это связано с тем, что повторяющиеся последовательности могут вызвать проскальзывание репликации, при котором цепь шаблона и ее копия сдвигают свои относительные положения, так что часть шаблона либо копируется дважды, либо пропускается.В результате новый полинуклеотид имеет соответственно большее или меньшее количество повторяющихся единиц (). Это основная причина, по которой микросателлитные последовательности настолько изменчивы, проскальзывание репликации иногда приводит к появлению нового варианта длины, добавляя к набору аллелей, уже присутствующих в популяции.
Рисунок 14.5
Проскальзывание репликации. На диаграмме показана репликация микросателлитного повтора CA из пяти единиц. Проскальзывание произошло во время репликации родительской молекулы, вставив дополнительную повторяющуюся единицу во вновь синтезированный полинуклеотид одной из дочерних (больше…)
Проскальзывание произошло во время репликации родительской молекулы, вставив дополнительную повторяющуюся единицу во вновь синтезированный полинуклеотид одной из дочерних (больше…)
Проскальзывание репликации, вероятно, также является причиной заболеваний, связанных с экспансией тринуклеотидных повторов, которые были обнаружены у людей в последние годы (Ashley and Warren, 1995). Каждое из этих нейродегенеративных заболеваний вызвано относительно коротким рядом тринуклеотидных повторов, которые удлиняются в два или более раз по сравнению с нормальной длиной. Например, человеческий ген HD содержит последовательность 5′-CAG-3′, повторяющуюся последовательно от 6 до 35 раз, кодирующую ряд глутаминов в белковом продукте.При болезни Гентингтона этот повтор расширяется до количества копий 36–121, увеличивая длину полиглутаминового тракта и приводя к дисфункциональному белку (Perutz, 1999). Некоторые другие заболевания человека также вызываются экспансией кодонов полиглутамина (4). Некоторые заболевания, связанные с умственной отсталостью, возникают в результате экспансии тринуклеотидов в лидерной области гена, что приводит к хрупкому участку, положению, в котором хромосома может сломаться (Sutherland et al. ., 1998). Также известны экспансии с участием интронных и концевых областей.
Некоторые заболевания, связанные с умственной отсталостью, возникают в результате экспансии тринуклеотидов в лидерной области гена, что приводит к хрупкому участку, положению, в котором хромосома может сломаться (Sutherland et al. ., 1998). Также известны экспансии с участием интронных и концевых областей.
Таблица 14.1
Примеры экспансии тринуклеотидных повторов человека.
Как генерируются триплетные расширения, точно не известно. Размер вставки намного больше, чем происходит при нормальном проскальзывании репликации, например, наблюдаемом с микросателлитными последовательностями, и как только расширение достигает определенной длины, оно, по-видимому, становится восприимчивым к дальнейшему расширению в последующих раундах репликации, так что болезнь становится становится все более тяжелым в последующих поколениях.Возможность того, что экспансия включает в себя образование шпилечных петель в ДНК, была выдвинута на основании наблюдения, что только ограниченное число тринуклеотидных последовательностей, как известно, подвергается экспансии, и все эти последовательности богаты GC и поэтому могут образовывать стабильные вторичные структуры. . Имеются также данные о том, что по крайней мере одна область триплетной экспансии — при атаксии Фридрейха — может образовывать структуру тройной спирали (Gacy et al. ., 1998). Исследования подобных экспансий триплетов у дрожжей показали, что они более распространены, когда ген RAD27 инактивирован (Freudenreich et al ., 1998), интересное наблюдение, поскольку RAD27 представляет собой дрожжевую версию гена млекопитающих FEN1, белка, участвующего в процессинге фрагментов Окадзаки (раздел 13.2.2). Это может указывать на то, что экспансия тринуклеотидных повторов вызвана аберрацией в синтезе отстающих нитей.
. Имеются также данные о том, что по крайней мере одна область триплетной экспансии — при атаксии Фридрейха — может образовывать структуру тройной спирали (Gacy et al. ., 1998). Исследования подобных экспансий триплетов у дрожжей показали, что они более распространены, когда ген RAD27 инактивирован (Freudenreich et al ., 1998), интересное наблюдение, поскольку RAD27 представляет собой дрожжевую версию гена млекопитающих FEN1, белка, участвующего в процессинге фрагментов Окадзаки (раздел 13.2.2). Это может указывать на то, что экспансия тринуклеотидных повторов вызвана аберрацией в синтезе отстающих нитей.
Мутации также вызываются химическими и физическими мутагенами
Многие химические вещества, которые естественным образом встречаются в окружающей среде, обладают мутагенными свойствами, и в последние годы они были дополнены другими химическими мутагенами, возникающими в результате производственной деятельности человека.Физические агенты, такие как радиация, также являются мутагенными.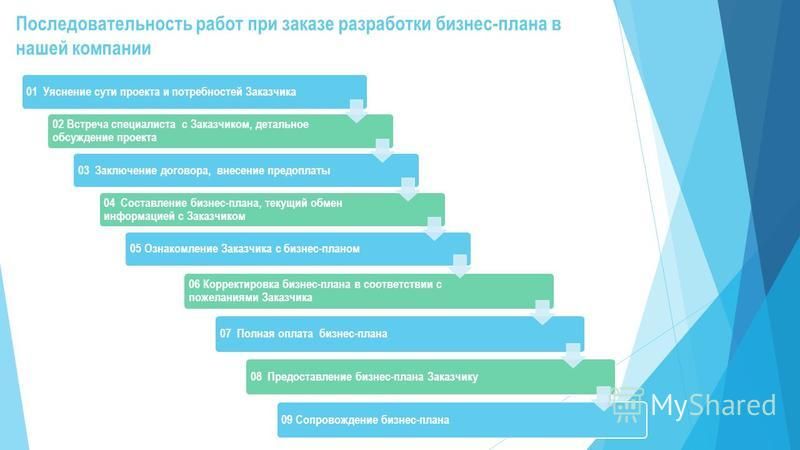 Большинство организмов подвергаются воздействию большего или меньшего количества этих различных мутагенов, в результате чего их геномы повреждаются.
Большинство организмов подвергаются воздействию большего или меньшего количества этих различных мутагенов, в результате чего их геномы повреждаются.
Термин «мутаген» определяется как химический или физический агент, вызывающий мутации . Это определение важно, потому что оно отличает мутагены от других типов агентов окружающей среды, которые вызывают повреждение клеток не путем мутаций (14). Между этими категориями есть совпадения (например, некоторые мутагены также являются канцерогенами), но каждый тип агента имеет различное биологическое действие.Определение «мутаген» также проводит различие между истинными мутагенами и другими агентами, которые повреждают ДНК, не вызывая мутаций, например, вызывая разрывы в молекулах ДНК. Этот тип повреждения может блокировать репликацию и вызывать гибель клетки, но это не мутация в строгом смысле этого слова, и поэтому возбудители не являются мутагенами.
Таблица 14.2
Категории агентов окружающей среды, вызывающих повреждение живых клеток.
Мутагены вызывают мутации тремя различными способами:
- Некоторые действуют как аналоги оснований и ошибочно используются в качестве субстратов при синтезе новой ДНК на репликационной вилке.
- Некоторые реагируют непосредственно с ДНК, вызывая структурные изменения, которые приводят к неправильному копированию цепи матрицы при репликации ДНК. Эти структурные изменения разнообразны, как мы увидим, когда рассмотрим отдельные мутагены.
- Некоторые мутагены косвенно воздействуют на ДНК. Сами по себе они не влияют на структуру ДНК, а заставляют клетку синтезировать такие химические вещества, как пероксиды, обладающие прямым мутагенным эффектом.
Спектр мутагенов настолько широк, что трудно разработать всеобъемлющую классификацию.Поэтому мы ограничим наше исследование наиболее распространенными типами. Для химических мутагенов они следующие:
- Аналоги оснований представляют собой пуриновые и пиримидиновые основания, которые достаточно похожи на стандартные основания, чтобы встраиваться в нуклеотиды, когда они синтезируются клеткой. Полученные необычные нуклеотиды затем можно использовать в качестве субстратов для синтеза ДНК во время репликации генома. Например, 5-бромурацил ( 5-bU 😉 обладает теми же свойствами спаривания оснований, что и тимин, и нуклеотиды, содержащие это основание, могут быть добавлены к дочернему полинуклеотиду в положениях, противоположных As в матрице.Мутагенный эффект возникает из-за того, что равновесие между двумя таутомерами 5-bU больше смещено в сторону более редкой формы енола , чем в случае тимина. Это означает, что во время следующего раунда репликации существует относительно высокая вероятность встречи полимеразы с енолом -5bU, который (как и енол -тимин) спаривается с G, а не с A (). Это приводит к точечной мутации (). 2-Аминопурин действует аналогичным образом: это аналог аденина с амино -таутомером, который спаривается с тимином, и имино таутомер, который спаривается с цитозином, причем форма имино встречается чаще, чем имино. -аденин и, следовательно, вызывает переходы T-C в ходе репликации ДНК.
- Дезаминирующие агенты также вызывают точечные мутации. Определенное количество оснований дезаминирования (удаление аминогруппы) происходит спонтанно в молекулах геномной ДНК, причем скорость увеличивается под действием таких химических веществ, как азотистая кислота, которая дезаминирует аденин, цитозин и гуанин (тимин не имеет аминогруппы и поэтому не может быть дезаминирован). ) и бисульфит натрия, который действует только на цитозин. Дезаминирование гуанина не является мутагенным, поскольку образующееся основание, ксантин, блокирует репликацию, когда оно появляется в матричном полинуклеотиде.Дезаминирование аденина дает гипоксантин (), который сочетается с С, а не с Т, а дезаминирование цитозина дает урацил, который сочетается с А, а не с G. Таким образом, дезаминирование этих двух оснований приводит к точковым мутациям при копировании матричной цепи.
- Алкилирующие агенты представляют собой третий тип мутагена, который может вызывать точечные мутации. Химические вещества, такие как этилметансульфонат ( EMS ) и диметилнитрозамин добавляют алкильные группы к нуклеотидам в молекулах ДНК, как и метилирующие агенты, такие как метилгалогениды, присутствующие в атмосфере, и продукты метаболизма нитритов.Эффект алкилирования зависит от положения, в котором модифицируется нуклеотид, и типа добавляемой алкильной группы. Метилирование, например, часто приводит к модифицированным нуклеотидам с измененными свойствами спаривания оснований и, таким образом, к точечным мутациям. Другие алкилирования блокируют репликацию, образуя поперечные связи между двумя цепями молекулы ДНК или добавляя большие алкильные группы, которые предотвращают развитие репликационного комплекса.
- Интеркалирующие агенты обычно связаны с инсерционными мутациями.Наиболее известным мутагеном этого типа является бромид этидия, который флуоресцирует при воздействии УФ-излучения и поэтому используется для выявления положения полос ДНК после электрофореза в агарозном геле (см. техническое примечание 2.1). Бромид этидия и другие интеркалирующие агенты представляют собой плоские молекулы, которые могут проскальзывать между парами оснований в двойной спирали, слегка раскручивая спираль и, следовательно, увеличивая расстояние между соседними парами оснований.
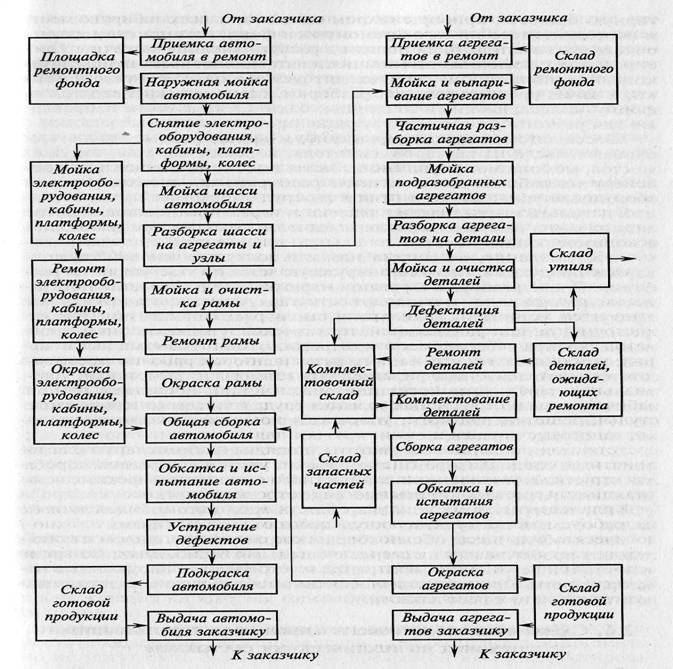 Полученные необычные нуклеотиды затем можно использовать в качестве субстратов для синтеза ДНК во время репликации генома. Например, 5-бромурацил ( 5-bU 😉 обладает теми же свойствами спаривания оснований, что и тимин, и нуклеотиды, содержащие это основание, могут быть добавлены к дочернему полинуклеотиду в положениях, противоположных As в матрице.Мутагенный эффект возникает из-за того, что равновесие между двумя таутомерами 5-bU больше смещено в сторону более редкой формы енола , чем в случае тимина. Это означает, что во время следующего раунда репликации существует относительно высокая вероятность встречи полимеразы с енолом -5bU, который (как и енол -тимин) спаривается с G, а не с A (). Это приводит к точечной мутации (). 2-Аминопурин действует аналогичным образом: это аналог аденина с амино -таутомером, который спаривается с тимином, и имино таутомер, который спаривается с цитозином, причем форма имино встречается чаще, чем имино.
Полученные необычные нуклеотиды затем можно использовать в качестве субстратов для синтеза ДНК во время репликации генома. Например, 5-бромурацил ( 5-bU 😉 обладает теми же свойствами спаривания оснований, что и тимин, и нуклеотиды, содержащие это основание, могут быть добавлены к дочернему полинуклеотиду в положениях, противоположных As в матрице.Мутагенный эффект возникает из-за того, что равновесие между двумя таутомерами 5-bU больше смещено в сторону более редкой формы енола , чем в случае тимина. Это означает, что во время следующего раунда репликации существует относительно высокая вероятность встречи полимеразы с енолом -5bU, который (как и енол -тимин) спаривается с G, а не с A (). Это приводит к точечной мутации (). 2-Аминопурин действует аналогичным образом: это аналог аденина с амино -таутомером, который спаривается с тимином, и имино таутомер, который спаривается с цитозином, причем форма имино встречается чаще, чем имино. -аденин и, следовательно, вызывает переходы T-C в ходе репликации ДНК.
-аденин и, следовательно, вызывает переходы T-C в ходе репликации ДНК. Химические вещества, такие как этилметансульфонат ( EMS ) и диметилнитрозамин добавляют алкильные группы к нуклеотидам в молекулах ДНК, как и метилирующие агенты, такие как метилгалогениды, присутствующие в атмосфере, и продукты метаболизма нитритов.Эффект алкилирования зависит от положения, в котором модифицируется нуклеотид, и типа добавляемой алкильной группы. Метилирование, например, часто приводит к модифицированным нуклеотидам с измененными свойствами спаривания оснований и, таким образом, к точечным мутациям. Другие алкилирования блокируют репликацию, образуя поперечные связи между двумя цепями молекулы ДНК или добавляя большие алкильные группы, которые предотвращают развитие репликационного комплекса.
Химические вещества, такие как этилметансульфонат ( EMS ) и диметилнитрозамин добавляют алкильные группы к нуклеотидам в молекулах ДНК, как и метилирующие агенты, такие как метилгалогениды, присутствующие в атмосфере, и продукты метаболизма нитритов.Эффект алкилирования зависит от положения, в котором модифицируется нуклеотид, и типа добавляемой алкильной группы. Метилирование, например, часто приводит к модифицированным нуклеотидам с измененными свойствами спаривания оснований и, таким образом, к точечным мутациям. Другие алкилирования блокируют репликацию, образуя поперечные связи между двумя цепями молекулы ДНК или добавляя большие алкильные группы, которые предотвращают развитие репликационного комплекса.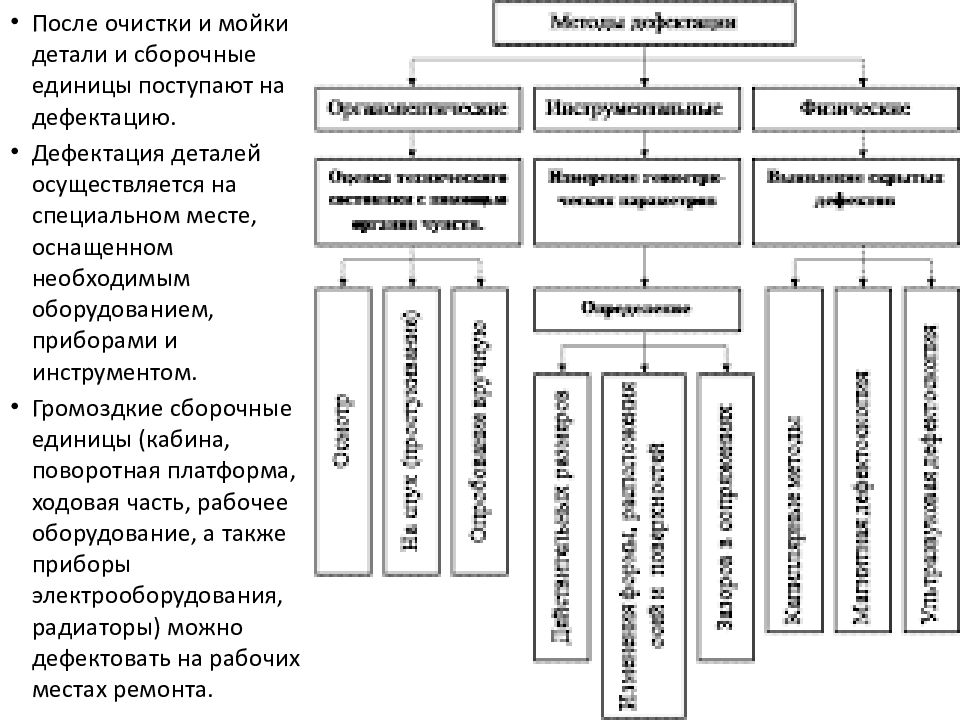 техническое примечание 2.1). Бромид этидия и другие интеркалирующие агенты представляют собой плоские молекулы, которые могут проскальзывать между парами оснований в двойной спирали, слегка раскручивая спираль и, следовательно, увеличивая расстояние между соседними парами оснований.
техническое примечание 2.1). Бромид этидия и другие интеркалирующие агенты представляют собой плоские молекулы, которые могут проскальзывать между парами оснований в двойной спирали, слегка раскручивая спираль и, следовательно, увеличивая расстояние между соседними парами оснований.Рисунок 14.6
5-Бромурацил и его мутагенное действие.См текст для деталей.
Рисунок 14.7
Гипоксантин представляет собой дезаминированную версию аденина. Нуклеозид, содержащий гипоксантин, называется инозином (см. табл. 10.5).
Рисунок 14.8
Мутагенный эффект этидия бромида. (A) Бромид этидия представляет собой плоскую пластинчатую молекулу, которая может располагаться между парами оснований двойной спирали. (B) Молекулы бромистого этидия показаны интеркалированными в спираль: молекулы видны (подробнее…)
Наиболее важными типами физического мутагена являются следующие:
- УФ-излучение с длиной волны 260 нм вызывает димеризацию соседних пиримидиновых оснований, особенно если они оба являются тиминами (), что приводит к образованию циклобутильного димера. Другие комбинации пиримидинов также образуют димеры, порядок частоты которых следующий: 5′-CT-3′ > 5′-TC-3′ > 5′-CC-3′. Пуриновые димеры встречаются гораздо реже. Индуцированная УФ-излучением димеризация обычно приводит к делеционным мутациям при копировании модифицированной цепи. Другим типом фотопродукта, индуцированного УФ-излучением, является повреждение (6-4), при котором атомы углерода номер 4 и 6 соседних пиримидинов становятся ковалентно связанными ().
- Ионизирующее излучение оказывает различное воздействие на ДНК в зависимости от типа излучения и его интенсивности.Могут возникать точечные, инсерционные и/или делеционные мутации, а также более тяжелые формы повреждения ДНК, препятствующие последующей репликации генома. Некоторые виды ионизирующего излучения воздействуют непосредственно на ДНК, другие воздействуют косвенно, стимулируя образование в клетке реактивных молекул, таких как перекиси.
- Тепло стимулирует вызванное водой расщепление β- N -гликозидной связи, которая соединяет основание с сахарным компонентом нуклеотида (). Это происходит чаще с пуринами, чем с пиримидинами, и приводит к AP (апуриновому/апиримидиновому) или безосновному сайту. Оставшийся сахар-фосфат нестабилен и быстро разлагается, оставляя разрыв, если молекула ДНК двухцепочечная (2). Эта реакция обычно не является мутагенной, поскольку в клетках имеются эффективные системы восстановления повреждений (раздел 14.2.1), что обнадеживает, если учесть, что в каждой клетке человека в день создается 10 000 АР-сайтов. Однако пробелы при определенных обстоятельствах приводят к мутациям, например, в E.coli при активации SOS-ответа, когда промежутки заполняются мышьяком независимо от идентичности нуклеотида в другой цепи (раздел 14.1.3).
 Другие комбинации пиримидинов также образуют димеры, порядок частоты которых следующий: 5′-CT-3′ > 5′-TC-3′ > 5′-CC-3′. Пуриновые димеры встречаются гораздо реже. Индуцированная УФ-излучением димеризация обычно приводит к делеционным мутациям при копировании модифицированной цепи. Другим типом фотопродукта, индуцированного УФ-излучением, является повреждение (6-4), при котором атомы углерода номер 4 и 6 соседних пиримидинов становятся ковалентно связанными ().
Другие комбинации пиримидинов также образуют димеры, порядок частоты которых следующий: 5′-CT-3′ > 5′-TC-3′ > 5′-CC-3′. Пуриновые димеры встречаются гораздо реже. Индуцированная УФ-излучением димеризация обычно приводит к делеционным мутациям при копировании модифицированной цепи. Другим типом фотопродукта, индуцированного УФ-излучением, является повреждение (6-4), при котором атомы углерода номер 4 и 6 соседних пиримидинов становятся ковалентно связанными (). Это происходит чаще с пуринами, чем с пиримидинами, и приводит к AP (апуриновому/апиримидиновому) или безосновному сайту. Оставшийся сахар-фосфат нестабилен и быстро разлагается, оставляя разрыв, если молекула ДНК двухцепочечная (2). Эта реакция обычно не является мутагенной, поскольку в клетках имеются эффективные системы восстановления повреждений (раздел 14.2.1), что обнадеживает, если учесть, что в каждой клетке человека в день создается 10 000 АР-сайтов. Однако пробелы при определенных обстоятельствах приводят к мутациям, например, в E.coli при активации SOS-ответа, когда промежутки заполняются мышьяком независимо от идентичности нуклеотида в другой цепи (раздел 14.1.3).
Это происходит чаще с пуринами, чем с пиримидинами, и приводит к AP (апуриновому/апиримидиновому) или безосновному сайту. Оставшийся сахар-фосфат нестабилен и быстро разлагается, оставляя разрыв, если молекула ДНК двухцепочечная (2). Эта реакция обычно не является мутагенной, поскольку в клетках имеются эффективные системы восстановления повреждений (раздел 14.2.1), что обнадеживает, если учесть, что в каждой клетке человека в день создается 10 000 АР-сайтов. Однако пробелы при определенных обстоятельствах приводят к мутациям, например, в E.coli при активации SOS-ответа, когда промежутки заполняются мышьяком независимо от идентичности нуклеотида в другой цепи (раздел 14.1.3).Рисунок 14.9
Фотопродукты, индуцированные УФ-облучением. Показан участок полинуклеотида, содержащий два соседних тиминовых основания. (A) Димер тимина содержит две ковалентные связи, индуцированные УФ-излучением: одна связывает атомы углерода в положении 6, а другая связывает атомы углерода (подробнее.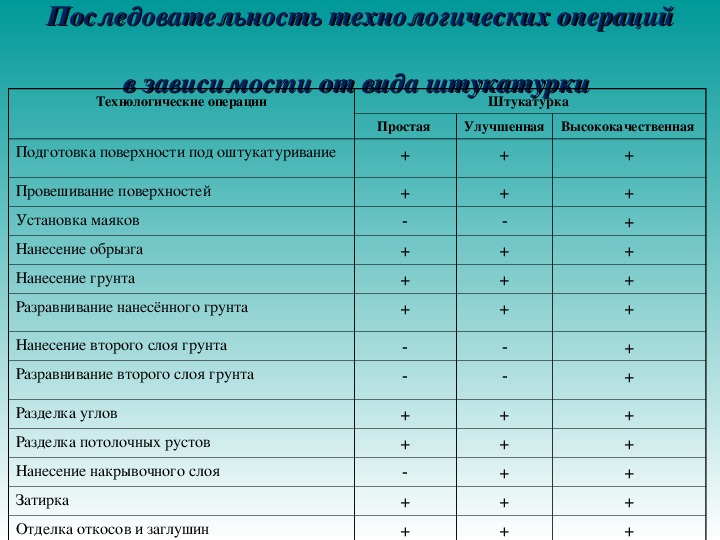 ..)
..)
Рисунок 14.10
Мутагенный эффект нагревания.(A) Нагрев вызывает гидролиз β- N -гликозидных связей, что приводит к образованию безосновного сайта в полинуклеотиде. (B) Схематическое изображение эффекта теплового гидролиза на молекулу двухцепочечной ДНК. Безосновательный (подробнее…)
14.1.2. Эффекты мутаций
При рассмотрении эффектов мутаций мы должны различать прямое влияние мутации на функционирование генома и ее косвенное влияние на фенотип организма, в котором она возникает.Прямой эффект относительно легко оценить, потому что мы можем использовать наше понимание структуры и экспрессии генов, чтобы предсказать влияние мутации на функцию генома. Косвенные эффекты более сложны, поскольку они относятся к фенотипу мутировавшего организма, который, как описано в разделе 7.2.2, часто трудно коррелировать с активностью отдельных генов.
Влияние мутаций на геномы
Многие мутации приводят к изменениям нуклеотидной последовательности, которые не влияют на функционирование генома. Эти молчащие мутации включают практически все те, которые встречаются в межгенной ДНК и в некодирующих компонентах генов и связанных с генами последовательностях. Другими словами, около 98,5% генома человека (см. вставку 1.4) можно мутировать без существенного эффекта.
Эти молчащие мутации включают практически все те, которые встречаются в межгенной ДНК и в некодирующих компонентах генов и связанных с генами последовательностях. Другими словами, около 98,5% генома человека (см. вставку 1.4) можно мутировать без существенного эффекта.
Мутации в кодирующих областях генов гораздо важнее. Во-первых, мы рассмотрим точечные мутации, которые изменяют последовательность триплетного кодона. Мутация этого типа будет иметь один из четырех эффектов ():
- Это может привести к синонимическому изменению, когда новый кодон определяет ту же аминокислоту, что и неизмененный кодон.Таким образом, синонимическое изменение является молчащей мутацией, поскольку оно не влияет на кодирующую функцию генома: мутировавший ген кодирует точно такой же белок, что и немутировавший ген.
- Это может привести к несинонимическому изменению, мутация изменяет кодон так, что он определяет другую аминокислоту. Таким образом, белок, кодируемый мутировавшим геном, имеет одну аминокислотную замену. Это часто не оказывает существенного влияния на биологическую активность белка, потому что большинство белков могут переносить по крайней мере несколько замен аминокислот без заметного влияния на их способность функционировать в клетке, но изменения некоторых аминокислот, например, в активной сайт фермента, имеют большее влияние.Несинонимическое изменение также называют мутацией missense .
- Мутация может преобразовать кодон, определяющий аминокислоту, в терминирующий кодон. Это бессмысленная мутация , и она приводит к укорочению белка, потому что трансляция мРНК останавливается на этом новом кодоне терминации, а не переходит к правильному кодону терминации ниже по течению. Влияние этого на активность белка зависит от того, какая часть полипептида потеряна: обычно эффект резкий, а белок нефункционален.
- Мутация может преобразовать терминирующий кодон в кодон, определяющий аминокислоту, что приводит к прочтению стоп-сигнала, поэтому белок удлиняется за счет дополнительной серии аминокислот на его С-конце. Большинство белков могут переносить короткие удлинения без влияния на функцию, но более длинные удлинения могут мешать сворачиванию белка и, таким образом, приводить к снижению активности.
 Это часто не оказывает существенного влияния на биологическую активность белка, потому что большинство белков могут переносить по крайней мере несколько замен аминокислот без заметного влияния на их способность функционировать в клетке, но изменения некоторых аминокислот, например, в активной сайт фермента, имеют большее влияние.Несинонимическое изменение также называют мутацией missense .
Это часто не оказывает существенного влияния на биологическую активность белка, потому что большинство белков могут переносить по крайней мере несколько замен аминокислот без заметного влияния на их способность функционировать в клетке, но изменения некоторых аминокислот, например, в активной сайт фермента, имеют большее влияние.Несинонимическое изменение также называют мутацией missense .Рисунок 14.11
Влияние точечных мутаций на кодирующую область гена.Показаны четыре различных эффекта точечных мутаций, как описано в тексте. Сквозная мутация приводит к тому, что ген выходит за пределы показанной здесь последовательности, лейцин (подробнее…)
Делеционные и инсерционные мутации также по-разному влияют на кодирующие способности генов (). Если количество удаленных или вставленных нуклеотидов равно трем или кратно трем, то один или несколько кодонов удаляются или добавляются, что приводит к потере или увеличению количества аминокислот, оказывающих различное влияние на функцию кодируемого белка.Делеции или вставки этого типа часто несущественны, но будут иметь значение, если, например, будут потеряны аминокислоты, участвующие в активном центре фермента, или если вставка нарушит важную вторичную структуру в белке. С другой стороны, если количество удаленных или вставленных нуклеотидов не равно трем или не кратно трем, тогда происходит сдвиг рамки считывания, когда все кодоны ниже мутации берутся из рамки считывания, отличной от той, которая используется в немутировавшем гене. Обычно это оказывает существенное влияние на функцию белка, поскольку большая или меньшая часть мутантного полипептида имеет совершенно другую последовательность, чем нормальный полипептид.
С другой стороны, если количество удаленных или вставленных нуклеотидов не равно трем или не кратно трем, тогда происходит сдвиг рамки считывания, когда все кодоны ниже мутации берутся из рамки считывания, отличной от той, которая используется в немутировавшем гене. Обычно это оказывает существенное влияние на функцию белка, поскольку большая или меньшая часть мутантного полипептида имеет совершенно другую последовательность, чем нормальный полипептид.
Труднее делать обобщения о влиянии мутаций, происходящих за пределами кодирующих областей генома. Любой сайт связывания белка восприимчив к точечным, инсерционным или делеционным мутациям, которые изменяют идентичность или относительное расположение нуклеотидов, участвующих во взаимодействии ДНК-белок. Таким образом, эти мутации могут инактивировать промоторы или регуляторные последовательности с предсказуемыми последствиями для экспрессии генов (разделы 9.2 и 9.3). Источники репликации, вероятно, могут стать нефункциональными из-за мутаций, которые изменяют, удаляют или нарушают последовательности, распознаваемые соответствующими связывающими белками (раздел 13. 2.1), но эти возможности плохо документированы. Также мало информации о потенциальном воздействии на экспрессию генов мутаций, влияющих на расположение нуклеосом (раздел 8.2.1).
2.1), но эти возможности плохо документированы. Также мало информации о потенциальном воздействии на экспрессию генов мутаций, влияющих на расположение нуклеосом (раздел 8.2.1).
Рисунок 14.13
Два возможных эффекта делеционных мутаций в области выше гена.
Одна из наиболее изученных областей касается мутаций, возникающих в интронах или на границах интрон-экзон. В этих регионах будут важны точечные мутации, если они изменяют нуклеотиды, участвующие во взаимодействиях РНК-белок и РНК-РНК, возникающих при сплайсинге различных типов интронов (разделы 10.1.3 и 10.2.3). Например, мутация G или T в ДНК-копии 5′-сайта сплайсинга интрона GU-AG или A или G в 3′-сайте сплайсинга нарушит сплайсинг, потому что правильная граница интрон-экзон больше не будет распознаваться. Это может означать, что интрон не удален из пре-мРНК, но более вероятно, что в качестве альтернативы будет использоваться криптический сайт сплайсинга (см. стр. 289). Также возможно, что мутация в интроне или экзоне создаст новый загадочный сайт, который предпочтительнее, чем настоящий сайт сплайсинга, который сам по себе не мутирован. Оба типа событий имеют одинаковый результат: перемещение активного сайта сплайсинга, ведущее к аберрантному сплайсингу. Это может привести к удалению части полученного белка, добавлению нового участка аминокислот или сдвигу рамки считывания. Несколько вариантов заболевания крови β-талассемия вызваны мутациями, которые приводят к скрытому выбору сайта сплайсинга во время процессинга транскриптов β-глобина.
Оба типа событий имеют одинаковый результат: перемещение активного сайта сплайсинга, ведущее к аберрантному сплайсингу. Это может привести к удалению части полученного белка, добавлению нового участка аминокислот или сдвигу рамки считывания. Несколько вариантов заболевания крови β-талассемия вызваны мутациями, которые приводят к скрытому выбору сайта сплайсинга во время процессинга транскриптов β-глобина.
Графа 14.1
Обнаружение мутаций. Быстрые процедуры обнаружения мутаций в молекулах ДНК. Многие генетические заболевания вызываются точечными мутациями, которые приводят к модификации или инактивации продукта гена.Методы обнаружения этих мутаций важны в двух (далее…)
Влияние мутаций на многоклеточные организмы
Теперь обратимся к косвенным эффектам, которые мутации оказывают на организмы, начиная с многоклеточных диплоидных эукариот, таких как человек. Первый вопрос, который следует рассмотреть, — это относительная важность одной и той же мутации в соматической клетке по сравнению с зародышевой клеткой. Поскольку соматические клетки не передают копии своих геномов следующему поколению, мутация соматической клетки важна только для организма, в котором она происходит: она не оказывает потенциального влияния на эволюцию.На самом деле, большинство мутаций соматических клеток не оказывают существенного влияния, даже если они приводят к гибели клеток, потому что в той же ткани есть много других идентичных клеток, и потеря одной клетки несущественна. Исключение составляет случай, когда мутация вызывает нарушение работы соматической клетки таким образом, что это вредно для организма, например, вызывая образование опухоли или другую раковую активность.
Поскольку соматические клетки не передают копии своих геномов следующему поколению, мутация соматической клетки важна только для организма, в котором она происходит: она не оказывает потенциального влияния на эволюцию.На самом деле, большинство мутаций соматических клеток не оказывают существенного влияния, даже если они приводят к гибели клеток, потому что в той же ткани есть много других идентичных клеток, и потеря одной клетки несущественна. Исключение составляет случай, когда мутация вызывает нарушение работы соматической клетки таким образом, что это вредно для организма, например, вызывая образование опухоли или другую раковую активность.
Мутации в зародышевых клетках более важны, потому что они могут быть переданы членам следующего поколения и затем будут присутствовать во всех клетках любого человека, который унаследует мутацию.Большинство мутаций, в том числе все молчащие и многие в кодирующих областях, по-прежнему не изменят фенотип организма каким-либо существенным образом. Те, которые действительно имеют эффект, можно разделить на две категории:
Те, которые действительно имеют эффект, можно разделить на две категории:
- Потеря функции является нормальным результатом мутации, которая снижает или отменяет активность белка. Большинство мутаций с потерей функции являются рецессивными (раздел 5.2.3), потому что у гетерозигот вторая копия хромосомы несет немутированную версию гена, кодирующего полностью функциональный белок, присутствие которого компенсирует эффект мутации (4).Есть некоторые исключения, когда мутация с потерей функции является доминирующей, одним из примеров является гаплонедостаточность, когда организм не может переносить примерно 50% снижение активности белка, от которого страдает гетерозигота. Это объясняет несколько генетических заболеваний у людей, в том числе синдром Марфана, который возникает в результате мутации гена белка соединительной ткани, называемого фибриллином.
Мутации с приобретением функции встречаются гораздо реже. Мутация должна быть такой, которая придает белку аномальную активность.
Многие мутации с приобретением функции находятся в регуляторных последовательностях, а не в кодирующих областях, и поэтому могут иметь ряд последствий. Например, мутация может привести к тому, что один или несколько генов будут экспрессироваться в неправильных тканях, и эти ткани приобретут функции, которых им обычно не хватает. В качестве альтернативы мутация может привести к сверхэкспрессии одного или нескольких генов, участвующих в контроле клеточного цикла, что приведет к неконтролируемому делению клеток и, следовательно, к раку. Из-за своей природы мутации с приобретением функции обычно являются доминирующими.
 Многие мутации с приобретением функции находятся в регуляторных последовательностях, а не в кодирующих областях, и поэтому могут иметь ряд последствий. Например, мутация может привести к тому, что один или несколько генов будут экспрессироваться в неправильных тканях, и эти ткани приобретут функции, которых им обычно не хватает. В качестве альтернативы мутация может привести к сверхэкспрессии одного или нескольких генов, участвующих в контроле клеточного цикла, что приведет к неконтролируемому делению клеток и, следовательно, к раку. Из-за своей природы мутации с приобретением функции обычно являются доминирующими.
Многие мутации с приобретением функции находятся в регуляторных последовательностях, а не в кодирующих областях, и поэтому могут иметь ряд последствий. Например, мутация может привести к тому, что один или несколько генов будут экспрессироваться в неправильных тканях, и эти ткани приобретут функции, которых им обычно не хватает. В качестве альтернативы мутация может привести к сверхэкспрессии одного или нескольких генов, участвующих в контроле клеточного цикла, что приведет к неконтролируемому делению клеток и, следовательно, к раку. Из-за своей природы мутации с приобретением функции обычно являются доминирующими.Рис. 14.14
Мутация с потерей функции обычно является рецессивной, поскольку функциональная версия гена присутствует во второй копии хромосомы.
Оценка влияния мутаций на фенотипы многоклеточных организмов может быть затруднена. Не все мутации оказывают немедленное воздействие: некоторые имеют отсроченное начало и придают измененный фенотип только позже в жизни человека. Другие проявляют непенетрантность у некоторых людей, никогда не проявляясь, даже если у человека есть доминантная мутация или он является гомозиготным рецессивным.У людей эти факторы усложняют попытки картирования мутаций, вызывающих заболевание, с помощью анализа родословной (раздел 5.2.4), потому что они вносят неопределенность в отношении того, какие члены родословной несут мутантный аллель.
Другие проявляют непенетрантность у некоторых людей, никогда не проявляясь, даже если у человека есть доминантная мутация или он является гомозиготным рецессивным.У людей эти факторы усложняют попытки картирования мутаций, вызывающих заболевание, с помощью анализа родословной (раздел 5.2.4), потому что они вносят неопределенность в отношении того, какие члены родословной несут мутантный аллель.
Воздействие мутаций на микроорганизмы
Мутации в микробах, таких как бактерии и дрожжи, также могут быть описаны как потеря или приобретение функции, но для микроорганизмов это не является ни нормальной, ни наиболее полезной схемой классификации. Вместо этого обычно предпринимают попытки более подробного описания фенотипа на основе свойств роста мутировавших клеток в различных культуральных средах.Это позволяет отнести большинство мутаций к одной из четырех категорий:
- Ауксотрофы — это клетки, которые будут расти только при наличии питательных веществ, которые не требуются немутировавшему организму. Например, E. coli обычно вырабатывает свой собственный триптофан благодаря ферментам, кодируемым пятью генами в триптофановом опероне (). Если один из этих генов мутирует таким образом, что его белковый продукт инактивируется, то клетка больше не способна производить триптофан и становится ауксотрофом по триптофану.Он не может выжить на среде, в которой отсутствует триптофан, и может расти только тогда, когда эта аминокислота предоставляется в качестве питательного вещества. Немутировавшие бактерии, не требующие дополнительных добавок в питательную среду, называются прототрофами.
- Условно-летальные мутанты не выдерживают определенных условий роста: в пермиссивных условиях они кажутся совершенно нормальными, но при переносе в рестриктивные условия проявляется мутантный фенотип. Термочувствительные мутанты являются типичными примерами условно-летальных мутантов.Чувствительные к температуре мутанты ведут себя как клетки дикого типа при низких температурах, но проявляют свой мутантный фенотип при повышении температуры выше определенного порога, который различен для каждого мутанта. Обычно это связано с тем, что мутация снижает стабильность белка, поэтому белок становится развернутым и, следовательно, неактивным при повышении температуры.
- Устойчивые к ингибиторам мутанты способны противостоять токсическому действию антибиотика или другого типа ингибитора.Существуют различные молекулярные объяснения этого типа мутантов. В некоторых случаях мутация изменяет структуру белка, на который нацелен ингибитор, так что последний больше не может связываться с белком и мешать его функции. На этом основана резистентность к стрептомицину E. coli , возникающая в результате изменения структуры рибосомного белка S12. Другая возможность заключается в том, что мутация изменяет свойства белка, ответственного за транспортировку ингибитора в клетку, что часто является способом приобретения устойчивости к токсичным металлам.
- Регуляторные мутанты имеют дефекты в промоторах и других регуляторных последовательностях. Эта категория включает конститутивных мутантов , которые непрерывно экспрессируют гены, которые в норме включаются и выключаются в различных условиях. Например, мутация в операторной последовательности лактозного оперона (раздел 9.3.1) может препятствовать связыванию репрессора и, таким образом, приводит к постоянной экспрессии лактозного оперона, даже когда лактоза отсутствует и гены должны быть выключены. ().
 Например, E. coli обычно вырабатывает свой собственный триптофан благодаря ферментам, кодируемым пятью генами в триптофановом опероне (). Если один из этих генов мутирует таким образом, что его белковый продукт инактивируется, то клетка больше не способна производить триптофан и становится ауксотрофом по триптофану.Он не может выжить на среде, в которой отсутствует триптофан, и может расти только тогда, когда эта аминокислота предоставляется в качестве питательного вещества. Немутировавшие бактерии, не требующие дополнительных добавок в питательную среду, называются прототрофами.
Например, E. coli обычно вырабатывает свой собственный триптофан благодаря ферментам, кодируемым пятью генами в триптофановом опероне (). Если один из этих генов мутирует таким образом, что его белковый продукт инактивируется, то клетка больше не способна производить триптофан и становится ауксотрофом по триптофану.Он не может выжить на среде, в которой отсутствует триптофан, и может расти только тогда, когда эта аминокислота предоставляется в качестве питательного вещества. Немутировавшие бактерии, не требующие дополнительных добавок в питательную среду, называются прототрофами. Обычно это связано с тем, что мутация снижает стабильность белка, поэтому белок становится развернутым и, следовательно, неактивным при повышении температуры.
Обычно это связано с тем, что мутация снижает стабильность белка, поэтому белок становится развернутым и, следовательно, неактивным при повышении температуры. Например, мутация в операторной последовательности лактозного оперона (раздел 9.3.1) может препятствовать связыванию репрессора и, таким образом, приводит к постоянной экспрессии лактозного оперона, даже когда лактоза отсутствует и гены должны быть выключены. ().
Например, мутация в операторной последовательности лактозного оперона (раздел 9.3.1) может препятствовать связыванию репрессора и, таким образом, приводит к постоянной экспрессии лактозного оперона, даже когда лактоза отсутствует и гены должны быть выключены. ().Рисунок 14.15
Ауксотрофный мутант по триптофану. Показаны две культуры в чашках Петри. Оба содержат минимальную среду, которая обеспечивает только основные потребности в питательных веществах для роста бактерий (азот, углерод и источники энергии, а также некоторые соли). Среда слева (подробнее…)
Рис. 14.16
Эффект конститутивной мутации в операторе лактозы. Последовательность оператора была изменена мутацией, и репрессор лактозы больше не может с ней связываться.В результате лактозный оперон транскрибируется все время, даже когда лактоза (подробнее…)
В дополнение к этим четырем категориям, многие мутации летальны и поэтому приводят к гибели мутантной клетки, в то время как другие не имеют никакого эффекта. . Последние менее распространены у микроорганизмов, чем у высших эукариот, потому что большинство микробных геномов относительно компактны, с небольшим количеством некодирующей ДНК. Мутации также могут быть негерметичными , что означает, что экспрессируется менее экстремальная форма мутантного фенотипа.Например, негерметичная версия триптофанового ауксотрофа, показанная на рис., будет медленно расти на минимальной среде, а не расти вообще.
. Последние менее распространены у микроорганизмов, чем у высших эукариот, потому что большинство микробных геномов относительно компактны, с небольшим количеством некодирующей ДНК. Мутации также могут быть негерметичными , что означает, что экспрессируется менее экстремальная форма мутантного фенотипа.Например, негерметичная версия триптофанового ауксотрофа, показанная на рис., будет медленно расти на минимальной среде, а не расти вообще.
14.1.3. Гипермутация и возможность запрограммированных мутаций
Могут ли клетки использовать мутации положительным образом, либо увеличивая скорость появления мутаций в своих геномах, либо направляя мутации в конкретные гены? На первый взгляд может показаться, что оба типа событий противоречат общепринятому мнению о том, что мутации происходят случайно, но, как мы увидим, гипермутации и запрограммированные мутации возможны, не противореча этой догме.
Гипермутация происходит, когда клетка позволяет увеличить скорость, с которой происходят мутации в ее геноме. Известно несколько примеров гипермутаций, один из которых является частью механизма, используемого позвоночными, включая человека, для создания разнообразных белков иммуноглобулинов. Мы уже касались этого явления в разделе 12.2.1, когда рассматривали перестройки генома, приводящие к соединению сегментов V, D, J и H генов тяжелой и легкой цепей иммуноглобулина (см. Ресурсы).Дополнительное разнообразие создается за счет гипермутации сегментов V-гена после сборки интактного гена иммуноглобулина (1), частота мутаций для этих сегментов на 6–7 порядков выше, чем частота фоновых мутаций, наблюдаемая в остальной части генома (Шеннон). и Вейгерт, 1998). Эта повышенная частота мутаций, по-видимому, является результатом необычного поведения системы восстановления несоответствия, которая обычно исправляет ошибки репликации. Во всех остальных положениях генома система репарации несоответствий исправляет ошибки репликации путем поиска несовпадений и замены нуклеотида в дочерней цепи, которая является только что синтезированной цепью и поэтому содержит ошибку (см.
Известно несколько примеров гипермутаций, один из которых является частью механизма, используемого позвоночными, включая человека, для создания разнообразных белков иммуноглобулинов. Мы уже касались этого явления в разделе 12.2.1, когда рассматривали перестройки генома, приводящие к соединению сегментов V, D, J и H генов тяжелой и легкой цепей иммуноглобулина (см. Ресурсы).Дополнительное разнообразие создается за счет гипермутации сегментов V-гена после сборки интактного гена иммуноглобулина (1), частота мутаций для этих сегментов на 6–7 порядков выше, чем частота фоновых мутаций, наблюдаемая в остальной части генома (Шеннон). и Вейгерт, 1998). Эта повышенная частота мутаций, по-видимому, является результатом необычного поведения системы восстановления несоответствия, которая обычно исправляет ошибки репликации. Во всех остальных положениях генома система репарации несоответствий исправляет ошибки репликации путем поиска несовпадений и замены нуклеотида в дочерней цепи, которая является только что синтезированной цепью и поэтому содержит ошибку (см. 2.3). В сегментах V-гена система репарации изменяет нуклеотид в родительской цепи и, таким образом, стабилизирует мутацию, а не исправляет ее (Cascalho et al. , 1998). Механизм, с помощью которого это достигается, еще не описан.
2.3). В сегментах V-гена система репарации изменяет нуклеотид в родительской цепи и, таким образом, стабилизирует мутацию, а не исправляет ее (Cascalho et al. , 1998). Механизм, с помощью которого это достигается, еще не описан.
Рисунок 14.17
Гипермутация сегмента V-гена интактного гена иммуноглобулина. См. рис. 12.15 для описания событий, ведущих к сборке гена иммуноглобулина.
Очевидное увеличение частоты мутаций, возникающее в результате модификации нормального процесса репарации ДНК, не противоречит догме о случайности мутаций.Однако возникли проблемы с отчетами, относящимися к 1988 г. (Cairns et al. ., 1988), в которых предполагалось, что E. coli способна направлять мутации в гены, мутация которых была бы полезной в условиях окружающей среды, в которых бактерия встречается. Первоначальные эксперименты включали штамм E. coli , который имел нонсенс-мутацию в опероне лактозы, инактивирующую белки, необходимые для утилизации этого сахара (Информация об исследованиях 14. 1).Бактерии рассеяли на агаризованной среде, в которой единственным источником углерода была лактоза. Это означало, что клетка могла расти и делиться только в том случае, если в опероне лактозы произошла вторая мутация, обращающая вспять эффекты бессмысленной мутации и, следовательно, позволяющая синтезировать ферменты лактозы. Мутации с таким эффектом происходили значительно чаще, чем ожидалось, и с большей скоростью, чем мутации в других частях геномов этих клеток E. coli .
1).Бактерии рассеяли на агаризованной среде, в которой единственным источником углерода была лактоза. Это означало, что клетка могла расти и делиться только в том случае, если в опероне лактозы произошла вторая мутация, обращающая вспять эффекты бессмысленной мутации и, следовательно, позволяющая синтезировать ферменты лактозы. Мутации с таким эффектом происходили значительно чаще, чем ожидалось, и с большей скоростью, чем мутации в других частях геномов этих клеток E. coli .
Графа 14.1
Запрограммированные мутации? В 1988 году были опубликованы поразительные результаты, предполагающие, что при некоторых обстоятельствах бактерии Escherichia coli способны целенаправленно мутировать, что позволяет клеткам адаптироваться к стрессу окружающей среды. Случайность мутаций (подробнее…)
Эти эксперименты показали, что бактерии могут программировать мутации в соответствии с давлением отбора, которому они подвергаются. Другими словами, окружающая среда может напрямую влиять на фенотип организма, как предполагал Ламарк, а не через случайные процессы, постулированные Дарвином. С такими радикальными последствиями неудивительно, что эксперименты долго обсуждались с многочисленными попытками обнаружить недостатки в их конструкции или альтернативные объяснения результатов. Вариации исходной экспериментальной системы позволили предположить, что результаты являются достоверными, и были описаны аналогичные события с другими бактериями. В настоящее время тестируются модели, основанные на амплификации генов, а не на селективных мутациях (Andersson et al. , 1998), и внимание также было направлено на возможную роль событий рекомбинации, таких как транспозиция вставочных элементов, в генерации запрограммированных мутаций (Foster , 1999).
С такими радикальными последствиями неудивительно, что эксперименты долго обсуждались с многочисленными попытками обнаружить недостатки в их конструкции или альтернативные объяснения результатов. Вариации исходной экспериментальной системы позволили предположить, что результаты являются достоверными, и были описаны аналогичные события с другими бактериями. В настоящее время тестируются модели, основанные на амплификации генов, а не на селективных мутациях (Andersson et al. , 1998), и внимание также было направлено на возможную роль событий рекомбинации, таких как транспозиция вставочных элементов, в генерации запрограммированных мутаций (Foster , 1999).
14.2. Восстановление ДНК
Принимая во внимание тысячи случаев повреждения генома каждый день, в сочетании с ошибками, возникающими при репликации генома, крайне важно, чтобы клетки обладали эффективными системами восстановления. Без этих систем репарации геном не смог бы поддерживать свои основные клеточные функции более чем за несколько часов до того, как ключевые гены будут инактивированы повреждением ДНК. Точно так же клеточные линии будут накапливать ошибки репликации с такой скоростью, что их геномы станут дисфункциональными после нескольких клеточных делений.
Точно так же клеточные линии будут накапливать ошибки репликации с такой скоростью, что их геномы станут дисфункциональными после нескольких клеточных делений.
Большинство клеток обладают четырьмя различными категориями системы репарации ДНК (; Lindahl and Wood, 1999):
Рисунок 14.18
Четыре категории систем репарации ДНК. См текст для деталей.
Большинство, если не все организмы, также обладают системами, которые позволяют им воспроизводить поврежденные участки своего генома без предварительного ремонта. Мы рассмотрим эти системы в разделе 14.2.5, а в разделе 14.2.6 мы рассмотрим болезни человека, возникающие в результате дефектов процессов репарации ДНК.
14.2.1. Системы прямой репарации заполняют разрывы и исправляют некоторые типы модификации нуклеотидов
Большинство типов повреждений ДНК, вызванных химическими или физическими мутагенами (раздел 14.1.1), можно устранить только путем удаления поврежденного нуклеотида с последующим ресинтезом новый участок ДНК, как показано на рис. Непосредственно восстановить можно только несколько типов поврежденных нуклеотидов:
Непосредственно восстановить можно только несколько типов поврежденных нуклеотидов:
- Разрывы можно восстановить с помощью ДНК-лигазы, если все, что произошло, это разрыв фосфодиэфирной связи без повреждения 5′-фосфатных и 3′-гидроксильных групп нуклеотидов по обе стороны от разрыва.Так часто бывает с зазубринами, возникающими в результате воздействия ионизирующего излучения.
- Некоторые формы повреждения алкилирования непосредственно обратимы с помощью ферментов, которые переносят алкильную группу с нуклеотида на собственные полипептидные цепи. Ферменты, способные это делать, известны во многих различных организмах и включают фермент Ada E. coli , который участвует в пассивном процессе, который эта бактерия способна активировать в ответ на повреждение ДНК.Ада удаляет алкильные группы, присоединенные к кислородным группам в положениях 4 и 6 тимина и гуанина соответственно, а также может восстанавливать метилированные фосфодиэфирные связи. Другие ферменты восстановления алкилирования имеют более ограниченную специфичность, например, человеческий MGMT (O 6 -метилгуанин-ДНК-метилтрансфераза), который, как следует из его названия, удаляет алкильные группы только из положения 6 гуанина.
- Циклобутиловые димеры восстанавливаются с помощью светозависимой прямой системы, называемой фотореактивацией.В E. coli процесс включает фермент, называемый ДНК-фотолиазой (более правильно называть дезоксирибодипиримидин-фотолиазой). При стимуляции светом с длиной волны от 300 до 500 нм фермент связывается с димерами циклобутил и превращает их обратно в исходные мономерные нуклеотиды. Фотореактивация является широко распространенным, но не универсальным типом репарации: она известна у многих, но не у всех бактерий, а также у многих эукариот, включая некоторых позвоночных, но отсутствует у человека и других плацентарных млекопитающих.Аналогичный тип фотореактивации включает фотолиазу (6-4) фотопродукта и приводит к репарации (6-4) повреждений. Ни E. coli , ни люди не имеют этого фермента, но им обладают многие другие организмы.

14.2.2. Эксцизионная репарация
Описанные выше прямые типы реверсирования повреждений важны, но они составляют очень второстепенный компонент механизмов репарации ДНК большинства организмов. Этот момент иллюстрируется черновыми последовательностями генома человека, которые, по-видимому, содержат только один ген, кодирующий белок, участвующий в прямой репарации (ген MGMT ), но которые имеют по крайней мере 40 генов для компонентов путей эксцизионной репарации. Вуд и др. ., 2001). Эти пути делятся на две категории:
Этот момент иллюстрируется черновыми последовательностями генома человека, которые, по-видимому, содержат только один ген, кодирующий белок, участвующий в прямой репарации (ген MGMT ), но которые имеют по крайней мере 40 генов для компонентов путей эксцизионной репарации. Вуд и др. ., 2001). Эти пути делятся на две категории:
- Иссечение основания Восстановление включает удаление поврежденного нуклеотидного основания, вырезание короткого фрагмента полинуклеотида вокруг созданного таким образом АР-сайта и ресинтез с помощью ДНК-полимеразы.
- Эксцизионная репарация нуклеотидов аналогична эксцизионной репарации оснований, но ей не предшествует удаление поврежденного основания, и она может воздействовать на более существенно поврежденные участки ДНК.
Мы рассмотрим каждый из этих путей по очереди.
Иссечение оснований восстанавливает многие типы поврежденных нуклеотидов
Иссечение оснований представляет собой наименее сложную из различных систем восстановления, которые включают удаление одного или нескольких поврежденных нуклеотидов с последующим ресинтезом ДНК для устранения образовавшегося разрыва. Он используется для репарации многих модифицированных нуклеотидов, основания которых подверглись относительно небольшим повреждениям в результате, например, воздействия алкилирующих агентов или ионизирующего излучения (раздел 14.1.1). Процесс инициируется ДНК-гликозилазой, которая расщепляет β- N -гликозидную связь между поврежденным основанием и сахарным компонентом нуклеотида ().Каждая ДНК-гликозилаза имеет ограниченную специфичность (), специфичность гликозилаз, которыми обладает клетка, определяет диапазон поврежденных нуклеотидов, которые могут быть репарированы путем удаления оснований. Большинство организмов способны справляться с дезаминированными основаниями, такими как урацил (дезаминированный цитозин) и гипоксантин (дезаминированный аденин), продуктами окисления, такими как 5-гидроксицитозин и тимингликоль, и метилированными основаниями, такими как 3-метиладенин, 7-метилгуанин и 2-метилцитозин. (Сиберг и др. ., 1995). Другие ДНК-гликозилазы удаляют нормальные основания как часть системы репарации несоответствия (раздел 14.
Он используется для репарации многих модифицированных нуклеотидов, основания которых подверглись относительно небольшим повреждениям в результате, например, воздействия алкилирующих агентов или ионизирующего излучения (раздел 14.1.1). Процесс инициируется ДНК-гликозилазой, которая расщепляет β- N -гликозидную связь между поврежденным основанием и сахарным компонентом нуклеотида ().Каждая ДНК-гликозилаза имеет ограниченную специфичность (), специфичность гликозилаз, которыми обладает клетка, определяет диапазон поврежденных нуклеотидов, которые могут быть репарированы путем удаления оснований. Большинство организмов способны справляться с дезаминированными основаниями, такими как урацил (дезаминированный цитозин) и гипоксантин (дезаминированный аденин), продуктами окисления, такими как 5-гидроксицитозин и тимингликоль, и метилированными основаниями, такими как 3-метиладенин, 7-метилгуанин и 2-метилцитозин. (Сиберг и др. ., 1995). Другие ДНК-гликозилазы удаляют нормальные основания как часть системы репарации несоответствия (раздел 14.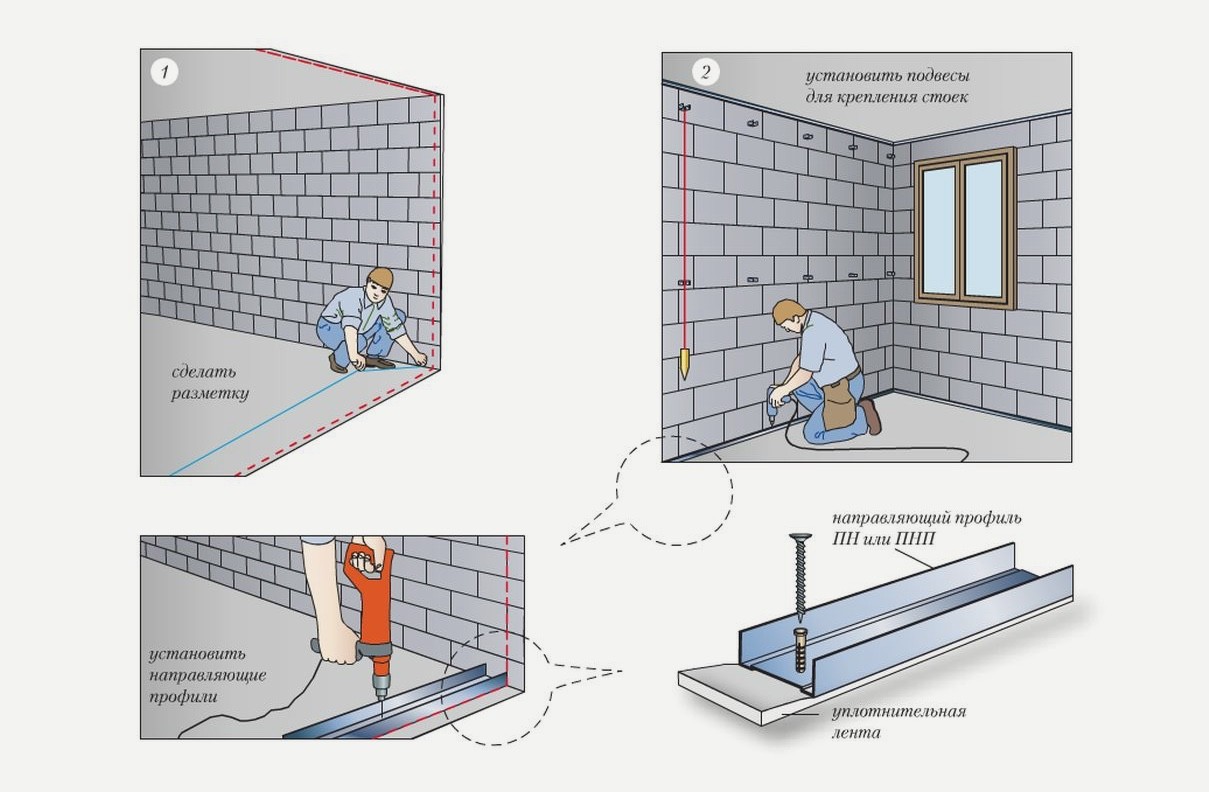 2.3). Считается, что большинство ДНК-гликозилаз, участвующих в эксцизионной репарации оснований, диффундируют вдоль малой бороздки двойной спирали ДНК в поисках поврежденных нуклеотидов, но некоторые из них могут быть связаны с ферментами репликации.
2.3). Считается, что большинство ДНК-гликозилаз, участвующих в эксцизионной репарации оснований, диффундируют вдоль малой бороздки двойной спирали ДНК в поисках поврежденных нуклеотидов, но некоторые из них могут быть связаны с ферментами репликации.
Рисунок 14.20
Иссечение основания. (A) Иссечение поврежденного нуклеотида ДНК-гликозилазой. (B) Схематическое изображение основного пути эксцизионной репарации.Альтернативные варианты пути описаны в тексте.
ДНК-гликозилаза удаляет поврежденное основание, «переворачивая» структуру в положение вне спирали и затем отсоединяя ее от полинуклеотида (Kunkel and Wilson, 1996; Roberts and Cheng, 1998). Это создает AP или безосновный сайт (см. ), который превращается в одиночный нуклеотидный пробел на втором этапе пути репарации (). Этот шаг может быть выполнен различными способами. В стандартном методе используется эндонуклеаза АР, такая как экзонуклеаза III или эндонуклеаза IV E.coli или APE1 человека, который разрезает фосфодиэфирную связь на 5′-стороне АР-сайта. Некоторые АР-эндонуклеазы могут также удалять сахар из АР-сайта, это все, что остается от поврежденного нуклеотида, но другие не обладают этой способностью и поэтому работают в сочетании с отдельной фосфодиэстеразой. Альтернативный путь превращения АР-сайта в разрыв использует эндонуклеазную активность, которой обладают некоторые ДНК-гликозилазы, которые могут сделать разрез на 3′-стороне АР-сайта, вероятно, в то же время, когда поврежденное основание удаляется, а затем снова путем удаления сахара фосфодиэстеразой.
Некоторые АР-эндонуклеазы могут также удалять сахар из АР-сайта, это все, что остается от поврежденного нуклеотида, но другие не обладают этой способностью и поэтому работают в сочетании с отдельной фосфодиэстеразой. Альтернативный путь превращения АР-сайта в разрыв использует эндонуклеазную активность, которой обладают некоторые ДНК-гликозилазы, которые могут сделать разрез на 3′-стороне АР-сайта, вероятно, в то же время, когда поврежденное основание удаляется, а затем снова путем удаления сахара фосфодиэстеразой.
Одиночный нуклеотидный пробел заполняется ДНК-полимеразой, используя спаривание оснований с неповрежденным основанием в другой цепи молекулы ДНК, чтобы гарантировать вставку правильного нуклеотида. У E. coli пробел заполняется ДНК-полимеразой I, а у млекопитающих — ДНК-полимеразой β (см. табл. 13.2; Sobol et al. ., 1996). Дрожжи, по-видимому, необычны тем, что они используют для этой цели свой основной ДНК-реплицирующий фермент, ДНК-полимеразу δ (Seeberg et al.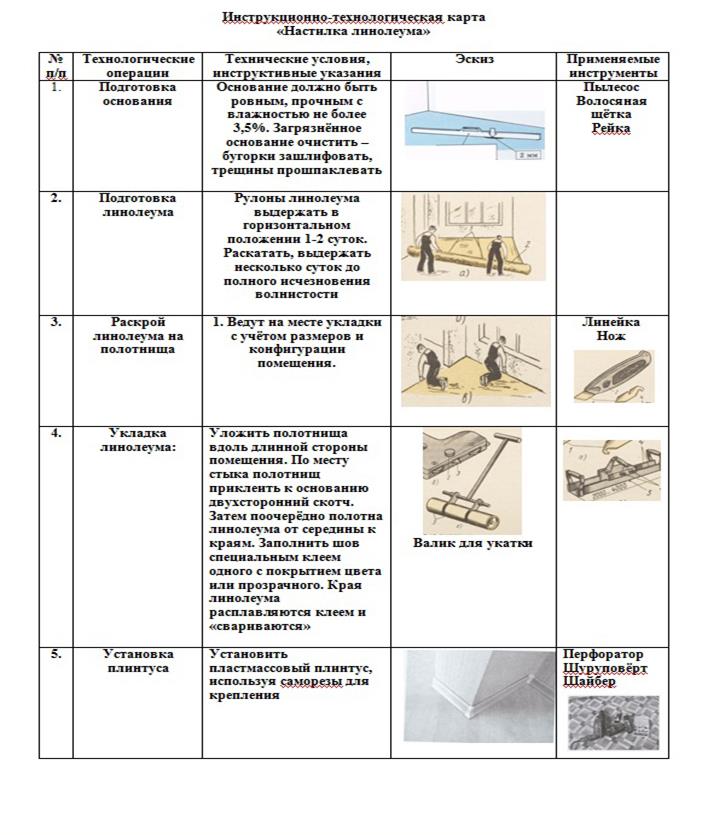 ., 1995).После заполнения промежутка последняя фосфодиэфирная связь устанавливается на место с помощью ДНК-лигазы.
., 1995).После заполнения промежутка последняя фосфодиэфирная связь устанавливается на место с помощью ДНК-лигазы.
Нуклеотидная эксцизионная репарация используется для коррекции более обширных типов повреждений
Нуклеотидная эксцизионная репарация имеет гораздо более широкую специфичность, чем система базового эксцизии, и способна воздействовать на более серьезные формы повреждений, такие как внутрицепочечные поперечные связи и модифицированные основания путем присоединения больших химических групп. Он также способен исправлять циклобутиловые димеры с помощью процесса восстановления в темноте, предоставляя тем организмам, у которых нет системы фотореактивации (например, людям), средства восстановления этого типа повреждений.
При эксцизионной репарации нуклеотидов сегмент одноцепочечной ДНК, содержащий поврежденный нуклеотид(ы), вырезается и заменяется новой ДНК. Таким образом, этот процесс подобен эксцизионной репарации оснований, за исключением того, что ему не предшествует селективное удаление оснований, и вырезается более длинный участок полинуклеотида. Наиболее изученным примером эксцизионной репарации нуклеотидов является процесс Short patch E. coli , названный так потому, что область полинуклеотида, которая вырезается и впоследствии «зашивается», относительно короткая, обычно 12 нуклеотидов в длину.
Наиболее изученным примером эксцизионной репарации нуклеотидов является процесс Short patch E. coli , названный так потому, что область полинуклеотида, которая вырезается и впоследствии «зашивается», относительно короткая, обычно 12 нуклеотидов в длину.
Репарация коротких участков инициируется мультиферментным комплексом, называемым эндонуклеазой UvrABC, иногда также называемой «эксинуклеазой». На первой стадии процесса к ДНК в месте повреждения присоединяется тример, состоящий из двух белков UvrA и одной копии UvrB. Как сайт распознается, неизвестно, но широкая специфичность процесса указывает на то, что отдельные типы повреждений не обнаруживаются напрямую и что комплекс должен искать более общий признак повреждения ДНК, такой как искажение двойной спирали.UvrA может быть частью комплекса, в наибольшей степени участвующей в локализации повреждения, потому что он диссоциирует, как только местонахождение найдено, и больше не играет никакой роли в процессе репарации. Уход UvrA позволяет UvrC связываться (), образуя димер UvrBC, который разрезает полинуклеотид по обе стороны от поврежденного сайта. Первый разрез производится UvrB по пятой фосфодиэфирной связи ниже поврежденного нуклеотида, а второй разрез производится UvrC по восьмой фосфодиэфирной связи выше по течению, что приводит к вырезанию 12 нуклеотидов, хотя есть некоторая вариабельность, особенно в положении места разреза UvrB.Вырезанный сегмент затем удаляется, обычно как неповрежденный олигонуклеотид, с помощью ДНК-хеликазы II, которая предположительно отщепляет сегмент, разрывая пары оснований, удерживающие его на второй цепи. UvrC также отделяется на этой стадии, но UvrB остается на месте и перекрывает зазор, образовавшийся в результате иссечения. Считается, что связанный UvrB предотвращает спаривание одноцепочечной области, которая подверглась воздействию, с самой собой, но альтернативные роли могут заключаться в предотвращении повреждения этой цепи или, возможно, в направлении ДНК-полимеразы к месту, которое необходимо отремонтировано.
Уход UvrA позволяет UvrC связываться (), образуя димер UvrBC, который разрезает полинуклеотид по обе стороны от поврежденного сайта. Первый разрез производится UvrB по пятой фосфодиэфирной связи ниже поврежденного нуклеотида, а второй разрез производится UvrC по восьмой фосфодиэфирной связи выше по течению, что приводит к вырезанию 12 нуклеотидов, хотя есть некоторая вариабельность, особенно в положении места разреза UvrB.Вырезанный сегмент затем удаляется, обычно как неповрежденный олигонуклеотид, с помощью ДНК-хеликазы II, которая предположительно отщепляет сегмент, разрывая пары оснований, удерживающие его на второй цепи. UvrC также отделяется на этой стадии, но UvrB остается на месте и перекрывает зазор, образовавшийся в результате иссечения. Считается, что связанный UvrB предотвращает спаривание одноцепочечной области, которая подверглась воздействию, с самой собой, но альтернативные роли могут заключаться в предотвращении повреждения этой цепи или, возможно, в направлении ДНК-полимеразы к месту, которое необходимо отремонтировано. Как и при эксцизионной репарации оснований, разрыв заполняется ДНК-полимеразой I, а последняя фосфодиэфирная связь синтезируется ДНК-лигазой.
Как и при эксцизионной репарации оснований, разрыв заполняется ДНК-полимеразой I, а последняя фосфодиэфирная связь синтезируется ДНК-лигазой.
. Поврежденный нуклеотид искажает спираль, потому что считается, что это один из сигналов распознавания тримера UvrAB, который инициирует процесс короткого участка. См. текст для (подробнее…)
E. coli также имеет систему эксцизионной репарации длиной нуклеотидов , которая включает белки Uvr, но отличается тем, что вырезаемый фрагмент ДНК может иметь длину до 2 т.п.н.Репарация длинных участков менее изучена, и этот процесс не изучен в деталях, но предполагается, что он работает с более обширными формами повреждений, возможно, с областями, где группы нуклеотидов, а не только отдельные, были изменены. Процесс эксцизионной репарации эукариотических нуклеотидов также называют «длинным участком», но он приводит к замене только 24–29 нуклеотидов ДНК. На самом деле, у эукариот нет системы «коротких заплат», и это название используется для того, чтобы отличить процесс от эксцизионной репарации основания. Система более сложная, чем у E. coli , и соответствующие ферменты, по-видимому, не являются гомологами белков Uvr. У человека задействовано не менее 16 белков, при этом нижний разрез делается в том же положении, что и в E. coli — пятая фосфодиэфирная связь, — но с более удаленным верхним разрезом, что приводит к более длительному вырезанию. Оба разреза осуществляются эндонуклеазами, которые атакуют одноцепочечную ДНК именно в месте ее соединения с двухцепочечным участком, что указывает на то, что до того, как были сделаны разрезы, ДНК вокруг места повреждения была расплавлена, предположительно, под действием хеликазы (4).Эта активность обеспечивается, по крайней мере частично, TFIIH, одним из компонентов инициирующего комплекса РНК-полимеразы II (см. табл. 9.5). Сначала предполагалось, что TFIIH просто играет двойную роль в клетке, функционируя отдельно как при транскрипции, так и при репарации, но теперь считается, что существует более прямая связь между этими двумя процессами (Lehmann, 1995; Svejstrup et al ).
Система более сложная, чем у E. coli , и соответствующие ферменты, по-видимому, не являются гомологами белков Uvr. У человека задействовано не менее 16 белков, при этом нижний разрез делается в том же положении, что и в E. coli — пятая фосфодиэфирная связь, — но с более удаленным верхним разрезом, что приводит к более длительному вырезанию. Оба разреза осуществляются эндонуклеазами, которые атакуют одноцепочечную ДНК именно в месте ее соединения с двухцепочечным участком, что указывает на то, что до того, как были сделаны разрезы, ДНК вокруг места повреждения была расплавлена, предположительно, под действием хеликазы (4).Эта активность обеспечивается, по крайней мере частично, TFIIH, одним из компонентов инициирующего комплекса РНК-полимеразы II (см. табл. 9.5). Сначала предполагалось, что TFIIH просто играет двойную роль в клетке, функционируя отдельно как при транскрипции, так и при репарации, но теперь считается, что существует более прямая связь между этими двумя процессами (Lehmann, 1995; Svejstrup et al ). ., 1996). Эта точка зрения подтверждается открытием репарации, связанной с транскрипцией, которая восстанавливает некоторые формы повреждений в матричных цепях генов, которые активно транскрибируются.Первым обнаруженным типом репарации, связанной с транскрипцией, была модифицированная версия эксцизионной репарации нуклеотидов, но теперь известно, что репарация с эксцизией оснований также связана с транскрипцией (Cooper et al. ., 1997). Эти открытия не означают, что нетранскрибируемые участки генома не репарируются. Процессы эксцизионной репарации защищают от повреждения весь геном, но вполне логично, что должны существовать специальные механизмы, направляющие процессы на транскрибируемые гены.Цепи-матрицы этих генов содержат биологическую информацию генома, и поддержание их целостности должно быть наивысшим приоритетом для систем репарации.
., 1996). Эта точка зрения подтверждается открытием репарации, связанной с транскрипцией, которая восстанавливает некоторые формы повреждений в матричных цепях генов, которые активно транскрибируются.Первым обнаруженным типом репарации, связанной с транскрипцией, была модифицированная версия эксцизионной репарации нуклеотидов, но теперь известно, что репарация с эксцизией оснований также связана с транскрипцией (Cooper et al. ., 1997). Эти открытия не означают, что нетранскрибируемые участки генома не репарируются. Процессы эксцизионной репарации защищают от повреждения весь геном, но вполне логично, что должны существовать специальные механизмы, направляющие процессы на транскрибируемые гены.Цепи-матрицы этих генов содержат биологическую информацию генома, и поддержание их целостности должно быть наивысшим приоритетом для систем репарации.
Рисунок 14.22
Схема событий, происходящих во время эксцизионной репарации нуклеотидов у эукариот. Эндонуклеазы, удаляющие поврежденный участок, делают надрезы именно в месте соединения одноцепочечных и двухцепочечных участков молекулы ДНК. Поэтому ДНК (подробнее…)
Поэтому ДНК (подробнее…)
14.2.3. Репарация несоответствия: исправление ошибок репликации
Каждая из систем репарации, которые мы рассмотрели до сих пор, — прямая репарация, эксцизионная репарация и эксцизионная репарация нуклеотидов — распознают повреждения ДНК, вызванные мутагенами, и воздействуют на них.Это означает, что они ищут аномальные химические структуры, такие как модифицированные нуклеотиды, циклобутиловые димеры и внутрицепочечные сшивки. Однако они не могут исправить несовпадения, возникающие в результате ошибок репликации, потому что несовпадающий нуклеотид никоим образом не является аномальным, это просто A, C, G или T, которые были вставлены в неправильное положение. Поскольку эти нуклеотиды выглядят точно так же, как и любые другие нуклеотиды, система восстановления несоответствия, которая исправляет ошибки репликации, должна обнаруживать не сам несовпадающий нуклеотид, а отсутствие спаривания оснований между родительской и дочерней цепями.Обнаружив несоответствие, система репарации вырезает часть дочернего полинуклеотида и заполняет пробел, аналогично репарации путем эксцизии оснований и нуклеотидов.
Описанная выше схема оставляет без ответа один важный вопрос. Ремонт должен производиться в дочернем полинуклеотиде, потому что именно в этой вновь синтезированной цепи произошла ошибка; исходный полинуклеотид имеет правильную последовательность. Как процесс восстановления узнает, какая прядь какая? В Э.coli ответ состоит в том, что дочерняя цепь на этой стадии недостаточно метилирована и поэтому может быть отличена от исходного полинуклеотида, имеющего полный набор метильных групп. ДНК E. coli метилирована из-за активности ДНК-аденинметилазы ( Dam ), которая превращает аденины в 6-метиладенины в последовательности 5′-GATC-3′, и ДНК-цитозинметилазы ( Dcm ), который превращает цитозины в 5-метилцитозины в 5′-CCAGG-3′ и 5′-CCTGG-3′.Эти метилирования не являются мутагенными, модифицированные нуклеотиды имеют те же свойства спаривания оснований, что и немодифицированные версии. Существует задержка между репликацией ДНК и метилированием дочерней цепи, и именно в это время система репарации сканирует ДНК на наличие несоответствий и вносит необходимые исправления в недостаточно метилированную дочернюю цепь.
Рисунок 14.23
Метилирование вновь синтезированной ДНК в Escherichia coli не происходит сразу после репликации, что дает возможность белкам восстановления несоответствия распознавать дочерние цепи и исправлять ошибки репликации.
E. coli имеет по крайней мере три системы репарации несоответствия, называемые «длинный участок», «короткий участок» и «очень короткий участок», названия которых указывают на относительную длину вырезанных и ресинтезированных сегментов. Система длинных заплат заменяет до kb или более ДНК и требует белков MutH, MutL и MutS, а также геликазу II ДНК, которую мы встретили во время эксцизионной репарации нуклеотидов. MutS распознает несоответствие, а MutH различает две нити, связываясь с неметилированными последовательностями 5′-GATC-3′ (10).Роль MutL неясна, но он может координировать активность двух других белков, так что MutH связывается с последовательностями 5′-GATC-3′ только вблизи сайтов несоответствия, распознаваемых MutS. После связывания MutH разрезает фосфодиэфирную связь непосредственно перед G в последовательности метилирования, а ДНК-хеликаза II отщепляет одну цепь. По-видимому, не существует фермента, который разрезал бы цепь после несоответствия; вместо этого отсоединенная одноцепочечная область расщепляется экзонуклеазой, которая следует за геликазой и продолжается за пределами сайта несоответствия.Затем пробел заполняется ДНК-полимеразой I и ДНК-лигазой. Сходные события, как полагают, происходят во время короткой и очень короткой репарации несоответствия, разница заключается в специфичности белков, которые распознают несоответствие. Система короткой заплаты, которая приводит к вырезанию сегмента длиной менее 10 нуклеотидов, начинается, когда MutY распознает несоответствие AG или AC, а система очень короткой репарации исправляет несоответствие G-T, которое распознается эндонуклеазой Vsr.
После связывания MutH разрезает фосфодиэфирную связь непосредственно перед G в последовательности метилирования, а ДНК-хеликаза II отщепляет одну цепь. По-видимому, не существует фермента, который разрезал бы цепь после несоответствия; вместо этого отсоединенная одноцепочечная область расщепляется экзонуклеазой, которая следует за геликазой и продолжается за пределами сайта несоответствия.Затем пробел заполняется ДНК-полимеразой I и ДНК-лигазой. Сходные события, как полагают, происходят во время короткой и очень короткой репарации несоответствия, разница заключается в специфичности белков, которые распознают несоответствие. Система короткой заплаты, которая приводит к вырезанию сегмента длиной менее 10 нуклеотидов, начинается, когда MutY распознает несоответствие AG или AC, а система очень короткой репарации исправляет несоответствие G-T, которое распознается эндонуклеазой Vsr.
Рис. 14.24
Исправление несовпадения длинных участков в Escherichia coli . См текст для деталей.
Эукариоты имеют гомологи белков E. coli Mut, и их процессы репарации несоответствия, вероятно, работают сходным образом (Kolodner, 2000). Единственное отличие состоит в том, что метилирование не может быть методом, используемым для различения родительских и дочерних полинуклеотидов. Метилирование участвует в репарации несоответствий в клетках млекопитающих, но ДНК некоторых эукариот, включая плодовых мух и дрожжей, не сильно метилирована; поэтому считается, что эти организмы должны использовать другой метод.Возможности включают ассоциацию между ферментами репарации и репликационным комплексом, так что репарация связана с синтезом ДНК, или использование одноцепочечных связывающих белков, которые маркируют родительскую цепь.
14.2.4. Репарация двухцепочечных разрывов ДНК
Одноцепочечный разрыв в двухцепочечной молекуле ДНК, например, возникающий в процессе эксцизионной репарации оснований и нуклеотидов, а также при некоторых типах окислительного повреждения, не представляет для клетки критического состояния. проблема.Двойная спираль сохраняет свою общую целостность, и разрыв может быть репарирован с помощью матрично-зависимого синтеза ДНК. Двухцепочечный разрыв более серьезен, потому что он превращает исходную двойную спираль в два отдельных фрагмента, которые необходимо снова собрать вместе, чтобы восстановить разрыв (2). Два разорванных конца должны быть защищены от дальнейшей деградации, которая может привести к появлению делеционных мутаций в точке репарированного разрыва. Процессы репарации также должны обеспечивать соединение правильных концов: если в ядре есть две сломанные хромосомы, то правильные пары должны быть сведены вместе, чтобы восстановить исходные структуры.Экспериментальные исследования клеток мышей показывают, что достичь такого результата сложно, и если две хромосомы сломаны, то неправильное восстановление, приводящее к гибридным структурам, происходит относительно часто (Richardson and Jasin, 2000). Даже если сломана только одна хромосома, все еще существует вероятность того, что естественный конец хромосомы можно спутать с разрывом и выполненным неправильным восстановлением.
проблема.Двойная спираль сохраняет свою общую целостность, и разрыв может быть репарирован с помощью матрично-зависимого синтеза ДНК. Двухцепочечный разрыв более серьезен, потому что он превращает исходную двойную спираль в два отдельных фрагмента, которые необходимо снова собрать вместе, чтобы восстановить разрыв (2). Два разорванных конца должны быть защищены от дальнейшей деградации, которая может привести к появлению делеционных мутаций в точке репарированного разрыва. Процессы репарации также должны обеспечивать соединение правильных концов: если в ядре есть две сломанные хромосомы, то правильные пары должны быть сведены вместе, чтобы восстановить исходные структуры.Экспериментальные исследования клеток мышей показывают, что достичь такого результата сложно, и если две хромосомы сломаны, то неправильное восстановление, приводящее к гибридным структурам, происходит относительно часто (Richardson and Jasin, 2000). Даже если сломана только одна хромосома, все еще существует вероятность того, что естественный конец хромосомы можно спутать с разрывом и выполненным неправильным восстановлением. Этот тип ошибок известен, несмотря на наличие особых белков, связывающих теломеры, которые маркируют естественные концы хромосом (раздел 2.2.1).
Этот тип ошибок известен, несмотря на наличие особых белков, связывающих теломеры, которые маркируют естественные концы хромосом (раздел 2.2.1).
Рис. 14.25
Ремонт одно- и двухнитевых разрывов. (A) Одноцепочечный разрыв не нарушает целостность двойной спирали. Открытая одиночная цепь покрыта белками PARP1, которые защищают эту неповрежденную цепь от разрыва и предотвращают ее участие (подробнее…)
Двухцепочечные разрывы образуются под воздействием ионизирующего излучения и некоторых химических мутагенов, а также клеткой контролируемым образом во время событий рекомбинации, таких как перестройки генома, которые соединяют сегменты гена иммуноглобулина и сегменты гена Т-клеточного рецептора в В- и Т-лимфоцитах (Раздел 12.2.1). Прогресс в понимании системы репарации разрывов был стимулирован исследованиями мутантных клеточных линий человека, которые привели к идентификации различных наборов генов, участвующих в этом процессе (Critchlow and Jackson, 1998). Эти гены определяют многокомпонентный белковый комплекс, который направляет ДНК-лигазу на разрыв (). Комплекс включает белок Ku, состоящий из двух неидентичных субъединиц, который связывает концы ДНК по обе стороны от разрыва (Walker et al. , 2001).Ku связывается с ДНК в ассоциации с протеинкиназой DNA-PK CS , которая активирует третий белок, XRCC4, который взаимодействует с ДНК-лигазой IV млекопитающих, направляя этот белок репарации на двухцепочечный разрыв.
Комплекс включает белок Ku, состоящий из двух неидентичных субъединиц, который связывает концы ДНК по обе стороны от разрыва (Walker et al. , 2001).Ku связывается с ДНК в ассоциации с протеинкиназой DNA-PK CS , которая активирует третий белок, XRCC4, который взаимодействует с ДНК-лигазой IV млекопитающих, направляя этот белок репарации на двухцепочечный разрыв.
Рисунок 14.26
Негомологичное соединение концов (NHEJ) у человека. (А) Процесс ремонта. Дополнительные белки, не показанные на диаграмме, также участвуют в NHEJ. К ним относятся протеинкиназы ATM и ATR (раздел 13.3.2), основная роль которых может заключаться в том, чтобы сигнализировать клетке о (больше…)
Процесс восстановления называется негомологичным соединением концов ( NHEJ ), это название указывает на то, что нет необходимости в гомологии между двумя молекулами, чьи концы соединяются, в отличие от других механизмов соединения концов, которые мы встретимся при изучении рекомбинации в разделе 14. 3. NHEJ рассматривается как тип рекомбинации, потому что, помимо восстановления разрывов, его можно использовать для соединения молекул или фрагментов, которые ранее не были соединены, создавая новые комбинации.Вариант системы NHEJ, вероятно, используется при конструировании генов иммуноглобулинов и рецепторов Т-клеток, но детали, вероятно, будут отличаться, поскольку эти запрограммированные перестройки генома включают промежуточные структуры, такие как шпильки ДНК, которые не видны во время конструирования. восстановление разрывов ДНК в результате повреждения.
3. NHEJ рассматривается как тип рекомбинации, потому что, помимо восстановления разрывов, его можно использовать для соединения молекул или фрагментов, которые ранее не были соединены, создавая новые комбинации.Вариант системы NHEJ, вероятно, используется при конструировании генов иммуноглобулинов и рецепторов Т-клеток, но детали, вероятно, будут отличаться, поскольку эти запрограммированные перестройки генома включают промежуточные структуры, такие как шпильки ДНК, которые не видны во время конструирования. восстановление разрывов ДНК в результате повреждения.
14.2.5. Обход повреждения ДНК во время репликации генома
Если область генома сильно повреждена, то вполне вероятно, что процессы восстановления будут перегружены.Затем клетка сталкивается с жестким выбором между смертью или попыткой репликации поврежденной области, даже если эта репликация может быть подвержена ошибкам и привести к мутировавшим дочерним молекулам. Столкнувшись с таким выбором, клетки E. coli неизменно выбирают второй вариант, вызывая одну из нескольких экстренных процедур для обхода мест серьезного повреждения. Наиболее изученным из этих обходных процессов является SOS-ответ, который позволяет клетке реплицировать свою ДНК, даже если матричные полинуклеотиды содержат AP-сайты и/или циклобутиловые димеры и другие фотопродукты, возникающие в результате воздействия химических мутагенов или УФ-излучения, которые обычно блокируют или, по крайней мере, задержка репликационного комплекса.Обход этих сайтов требует создания мутасомы, состоящей из комплекса UmuD’ 2 C (также называемого ДНК-полимеразой V, тримера, состоящего из двух белков UmuD’ и одной копии UmuC) и нескольких копий белка RecA . (Гудман, 2000). Последний представляет собой одноцепочечный ДНК-связывающий белок, который покрывает поврежденные цепи, позволяя комплексу UmuD’ 2 C вытеснять ДНК-полимеразу III и осуществлять подверженный ошибкам синтез ДНК до тех пор, пока поврежденный участок не будет пройден и ДНК-полимераза III может взять на себя еще раз ().
Наиболее изученным из этих обходных процессов является SOS-ответ, который позволяет клетке реплицировать свою ДНК, даже если матричные полинуклеотиды содержат AP-сайты и/или циклобутиловые димеры и другие фотопродукты, возникающие в результате воздействия химических мутагенов или УФ-излучения, которые обычно блокируют или, по крайней мере, задержка репликационного комплекса.Обход этих сайтов требует создания мутасомы, состоящей из комплекса UmuD’ 2 C (также называемого ДНК-полимеразой V, тримера, состоящего из двух белков UmuD’ и одной копии UmuC) и нескольких копий белка RecA . (Гудман, 2000). Последний представляет собой одноцепочечный ДНК-связывающий белок, который покрывает поврежденные цепи, позволяя комплексу UmuD’ 2 C вытеснять ДНК-полимеразу III и осуществлять подверженный ошибкам синтез ДНК до тех пор, пока поврежденный участок не будет пройден и ДНК-полимераза III может взять на себя еще раз ().
Рисунок 14.27
Реакция SOS Escherichia coli . См текст для деталей.
См текст для деталей.
Реакция SOS в первую очередь рассматривается как последняя лучшая возможность для бактерии реплицировать свою ДНК и, следовательно, выжить в неблагоприятных условиях. Однако ценой выживания является повышенная частота мутаций, потому что мутасома не восстанавливает повреждения, а просто позволяет реплицироваться поврежденному участку полинуклеотида. Когда она сталкивается с поврежденным положением в матричной ДНК, полимераза выбирает нуклеотид более или менее случайным образом, хотя и с некоторым предпочтением для размещения А напротив АР-сайта: в результате частота ошибок в процессе репликации увеличивается.Было высказано предположение, что эта повышенная частота мутаций является целью SOS-реакции, поскольку мутация является в некотором роде благоприятной реакцией на повреждение ДНК, но эта идея остается спорной (Chicurel, 2001).
Некоторое время SOS-ответ считался единственным процессом обхода повреждения у бактерий, но теперь мы понимаем, что по крайней мере две другие полимеразы E.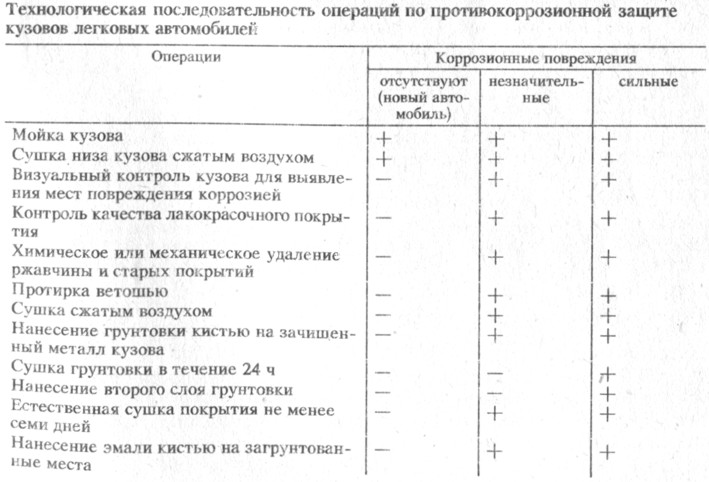 coli действуют аналогичным образом, хотя и с различными типами повреждений. Это ДНК-полимераза II, которая может обходить нуклеотиды, связанные с мутагенными химическими веществами, такими как N -2-ацетиламинофлуорен, и ДНК-полимераза IV (также называемая DinB), которая может реплицироваться через область матричной ДНК, в которой два родительских полинуклеотида имеют смещаются (Lindahl and Wood, 1999; Hanaoka, 2001).Обходные полимеразы также были обнаружены в эукариотических клетках. К ним относятся ДНК-полимераза η, которая может обходить циклобутиловые димеры (Johnson et al. ., 1999), и ДНК-полимеразы ι и ζ, которые работают вместе для репликации через фотопродукты и AP-сайты (Johnson et al. ., 2000).
coli действуют аналогичным образом, хотя и с различными типами повреждений. Это ДНК-полимераза II, которая может обходить нуклеотиды, связанные с мутагенными химическими веществами, такими как N -2-ацетиламинофлуорен, и ДНК-полимераза IV (также называемая DinB), которая может реплицироваться через область матричной ДНК, в которой два родительских полинуклеотида имеют смещаются (Lindahl and Wood, 1999; Hanaoka, 2001).Обходные полимеразы также были обнаружены в эукариотических клетках. К ним относятся ДНК-полимераза η, которая может обходить циклобутиловые димеры (Johnson et al. ., 1999), и ДНК-полимеразы ι и ζ, которые работают вместе для репликации через фотопродукты и AP-сайты (Johnson et al. ., 2000).
14.2.6. Дефекты репарации ДНК лежат в основе болезней человека, включая рак
Важность репарации ДНК подчеркивается количеством и тяжестью наследственных заболеваний человека, связанных с дефектами одного из процессов репарации.Одним из наиболее охарактеризованных из них является пигментная ксеродерма, которая возникает в результате мутации в любом из нескольких генов белков, участвующих в эксцизионной репарации нуклеотидов. Иссечение нуклеотидов — единственный способ, с помощью которого человеческие клетки могут восстанавливать циклобутиловые димеры и другие фотопродукты, поэтому неудивительно, что симптомы пигментной ксеродермы включают повышенную чувствительность к УФ-излучению, пациенты страдают от большего количества мутаций, чем обычно, под воздействием солнечного света, что часто приводит к рак кожи (Lehmann, 1995).Трихотиодистрофия также вызывается дефектами эксцизионной репарации нуклеотидов, но это более сложное заболевание, которое, хотя и не связано с раком, обычно включает проблемы как с кожей, так и с нервной системой.
Иссечение нуклеотидов — единственный способ, с помощью которого человеческие клетки могут восстанавливать циклобутиловые димеры и другие фотопродукты, поэтому неудивительно, что симптомы пигментной ксеродермы включают повышенную чувствительность к УФ-излучению, пациенты страдают от большего количества мутаций, чем обычно, под воздействием солнечного света, что часто приводит к рак кожи (Lehmann, 1995).Трихотиодистрофия также вызывается дефектами эксцизионной репарации нуклеотидов, но это более сложное заболевание, которое, хотя и не связано с раком, обычно включает проблемы как с кожей, так и с нервной системой.
Некоторые заболевания связаны с дефектами связанного с транскрипцией компонента эксцизионной репарации нуклеотидов. К ним относятся рак молочной железы и яичников, ген BRCA1 , который придает предрасположенность к этим видам рака, кодирующий белок, который, по крайней мере косвенно, участвует в репарации, связанной с транскрипцией (Gowen et al ., 1998) и синдром Коккейна, сложное заболевание, проявляющееся ростовыми и неврологическими нарушениями (Hanawalt, 2000). Недостаточность связанной с транскрипцией репарации также была выявлена у людей, страдающих синдромом предрасположенности к раку, называемым HNPCC (наследственный неполипозный колоректальный рак; Mellon et al. ., 1996), хотя первоначально это заболевание было идентифицировано как дефект в восстановление несоответствия (Колоднер, 1995). Атаксия-телеангиэктазия, симптомы которой включают чувствительность к ионизирующему излучению, возникает в результате дефектов в гене ATX , который участвует в процессе обнаружения повреждений (раздел 13.3.2). Другими заболеваниями, связанными с нарушением репарации ДНК, являются синдромы Блума и Вернера, которые вызываются инактивацией геликазы ДНК, которая может играть роль в NHEJ (Shen and Loeb, 2000; Wu and Hickson, 2001), и анемия Фанкони. , который придает чувствительность к химическим веществам, вызывающим перекрестные связи в ДНК, но биохимическая основа которых еще не известна.
Недостаточность связанной с транскрипцией репарации также была выявлена у людей, страдающих синдромом предрасположенности к раку, называемым HNPCC (наследственный неполипозный колоректальный рак; Mellon et al. ., 1996), хотя первоначально это заболевание было идентифицировано как дефект в восстановление несоответствия (Колоднер, 1995). Атаксия-телеангиэктазия, симптомы которой включают чувствительность к ионизирующему излучению, возникает в результате дефектов в гене ATX , который участвует в процессе обнаружения повреждений (раздел 13.3.2). Другими заболеваниями, связанными с нарушением репарации ДНК, являются синдромы Блума и Вернера, которые вызываются инактивацией геликазы ДНК, которая может играть роль в NHEJ (Shen and Loeb, 2000; Wu and Hickson, 2001), и анемия Фанкони. , который придает чувствительность к химическим веществам, вызывающим перекрестные связи в ДНК, но биохимическая основа которых еще не известна.
14.3. Рекомбинация
Без рекомбинации геномы были бы относительно статичными структурами, претерпевающими очень мало изменений.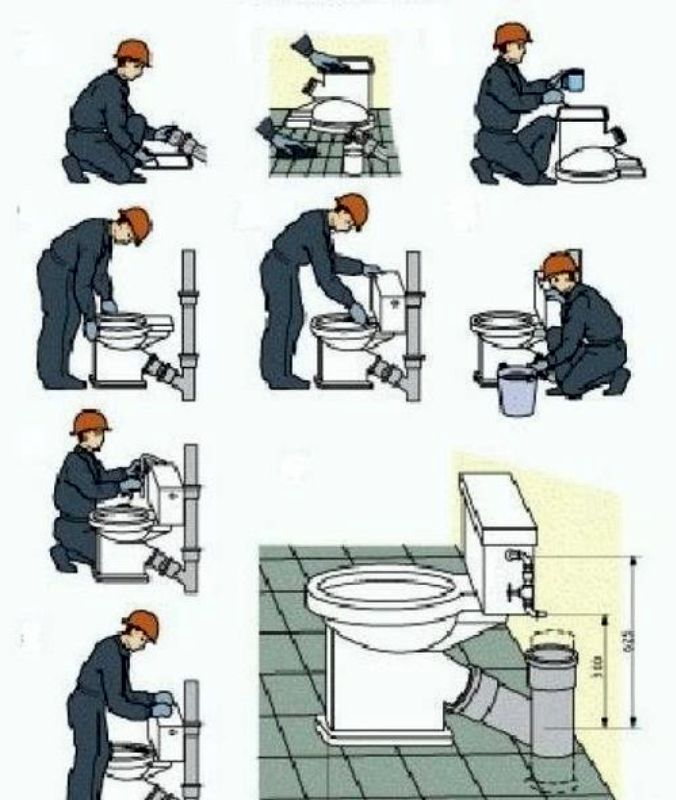 Постепенное накопление мутаций в течение длительного времени привело бы к мелкомасштабным изменениям в нуклеотидной последовательности генома, но не произошло бы более обширной перестройки, в которой играет роль рекомбинация, и эволюционный потенциал генома был бы снижен. быть строго ограничены.
Постепенное накопление мутаций в течение длительного времени привело бы к мелкомасштабным изменениям в нуклеотидной последовательности генома, но не произошло бы более обширной перестройки, в которой играет роль рекомбинация, и эволюционный потенциал генома был бы снижен. быть строго ограничены.
Рекомбинация была впервые признана как процесс, отвечающий за кроссинговер и обмен сегментами ДНК между гомологичными хромосомами во время мейоза эукариотических клеток (см. ), и впоследствии была вовлечена в интеграцию перенесенной ДНК в бактериальные геномы после конъюгации, трансдукции или трансформации (Раздел 5.2.4). Биологическое значение этих процессов стимулировало первые попытки описания молекулярных событий, участвующих в рекомбинации, и привело к модели Холлидея (Holliday, 1964), с которой мы начнем наше изучение рекомбинации.
14.3.1. Гомологическая рекомбинация
Модель Холлидея относится к типу рекомбинации, называемой вообще или гомологичной рекомбинации. Это наиболее важная версия рекомбинации в природе, отвечающая за мейотический кроссинговер и интеграцию перенесенной ДНК в бактериальные геномы.
Это наиболее важная версия рекомбинации в природе, отвечающая за мейотический кроссинговер и интеграцию перенесенной ДНК в бактериальные геномы.
Модель Холлидея для гомологичной рекомбинации
Модель Холлидея описывает рекомбинацию между двумя гомологичными двухцепочечными молекулами, молекулами с идентичными или почти идентичными последовательностями, но в равной степени применимыми к двум различным молекулам, которые имеют ограниченный участок гомологии, или к одному молекула, которая рекомбинирует сама с собой, поскольку содержит два отдельных участка, гомологичных друг другу.
Центральным элементом модели является образование гетеродуплекса в результате обмена полинуклеотидными сегментами между двумя гомологичными молекулами ().Первоначально гетеродуплекс стабилизируется за счет спаривания оснований между каждой перенесенной цепью и интактным полинуклеотидом молекулы-реципиента, причем такое спаривание оснований возможно из-за схожести последовательностей между двумя молекулами. Затем щели запечатываются ДНК-лигазой, что дает структуру Холлидея. Эта структура является динамической, миграция ветвей приводит к обмену более длинными сегментами ДНК, если две спирали вращаются в одном направлении.
Затем щели запечатываются ДНК-лигазой, что дает структуру Холлидея. Эта структура является динамической, миграция ветвей приводит к обмену более длинными сегментами ДНК, если две спирали вращаются в одном направлении.
Рисунок 14.28
Модель Холлидея для гомологичной рекомбинации.
Разделение или расщепление структуры Холлидея обратно на отдельные двухцепочечные молекулы происходит путем расщепления по точке ветвления. Это ключ ко всему процессу, потому что разрез может быть выполнен в любой из двух ориентаций, что становится очевидным при рассмотрении трехмерной конфигурации или формы ци структуры Холлидея (см. рис. 1). Эти два сокращения имеют очень разные результаты. Если сделать разрез влево-вправо поперек хи-формы, как показано на рис. , то все, что произойдет, это то, что короткий сегмент полинуклеотида, соответствующий расстоянию, пройденному ветвью структуры Холлидея, переместится между двумя молекулами.С другой стороны, разрез вверх-вниз приводит к взаимному обмену цепями, когда двухцепочечная ДНК переносится между двумя молекулами, так что конец одной молекулы заменяется на конец другой молекулы.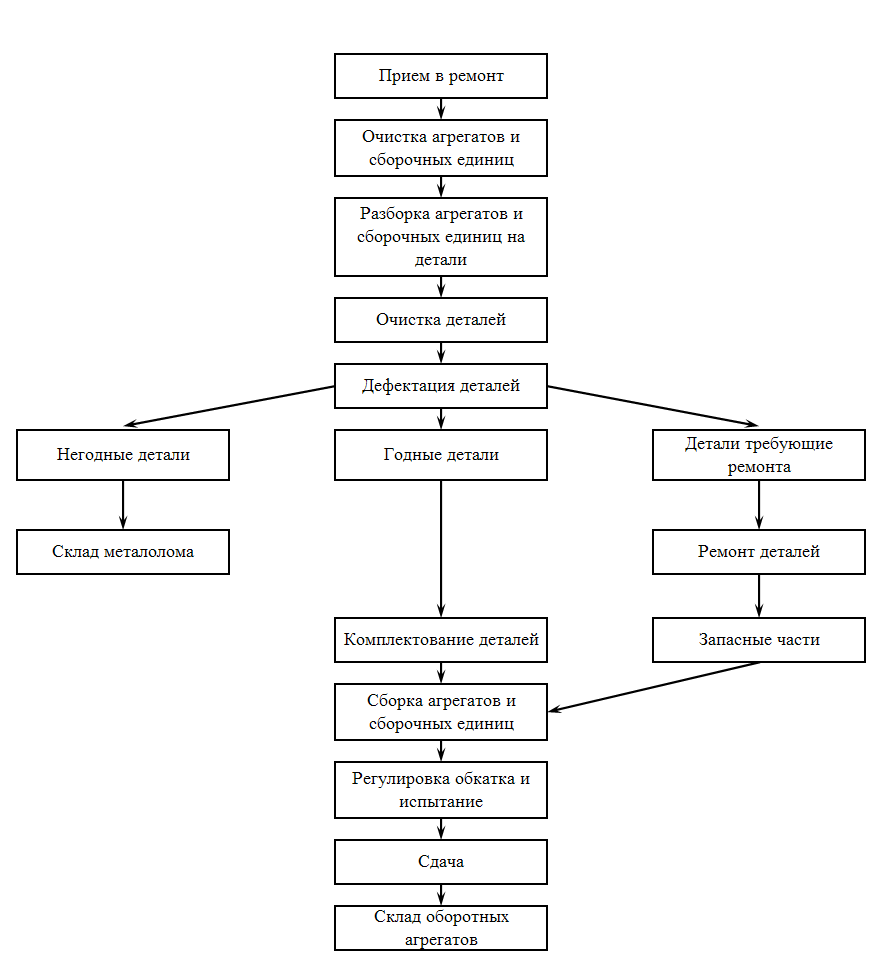 Это перенос ДНК, наблюдаемый при кроссинговере.
Это перенос ДНК, наблюдаемый при кроссинговере.
До сих пор мы игнорировали один аспект модели Холлидея. Так взаимодействуют две двухцепочечные молекулы в начале процесса образования гетеродуплекса. В исходной схеме две молекулы выстраивались друг относительно друга, и одноцепочечные разрывы появлялись в эквивалентных положениях каждой спирали.Это приводило к образованию свободных одноцепочечных концов, которые можно было обменивать, что приводило к гетеродуплексу. Эта особенность модели подверглась критике, потому что нельзя было предложить механизм, гарантирующий, что разрывы будут происходить точно в одном и том же месте на каждой молекуле. Модификация Мезельсона-Раддинга (Meselson and Radding, 1975) предлагает более удовлетворительную схему, согласно которой одноцепочечный разрыв происходит только в одной из двойных спиралей, а образовавшийся свободный конец «вторгается» в неразорванную двойную спираль в гомологичном положении и смещая одну из своих нитей, образуя D-петлю (). Последующее расщепление смещенной нити на стыке ее одноцепочечной и парной областей приводит к образованию гетеродуплекса.
Последующее расщепление смещенной нити на стыке ее одноцепочечной и парной областей приводит к образованию гетеродуплекса.
Рисунок 14.29
Две схемы инициации гомологичной рекомбинации. (A) Инициация, как описано в исходной модели гомологичной рекомбинации. (B) Модификация Мезельсона-Раддинга, которая предлагает более правдоподобную серию событий для образования гетеродуплекса. (подробнее…)
Белки, участвующие в гомологичной рекомбинации в
E.coli Модель Холлидея и модификация Мезельсона-Раддинга относятся к гомологичной рекомбинации во всех организмах, но, как и во многих областях молекулярной биологии, первоначальный прогресс в понимании того, как этот процесс осуществляется в клетке, был достигнут с E. coli . Исследуемая система специфической рекомбинации имеет кольцевой геном E. coli в качестве второго партнера и линейный фрагмент хромосомы в качестве второго партнера, что имеет место при конъюгации, трансдукции или трансформации бактериальных клеток (раздел 5.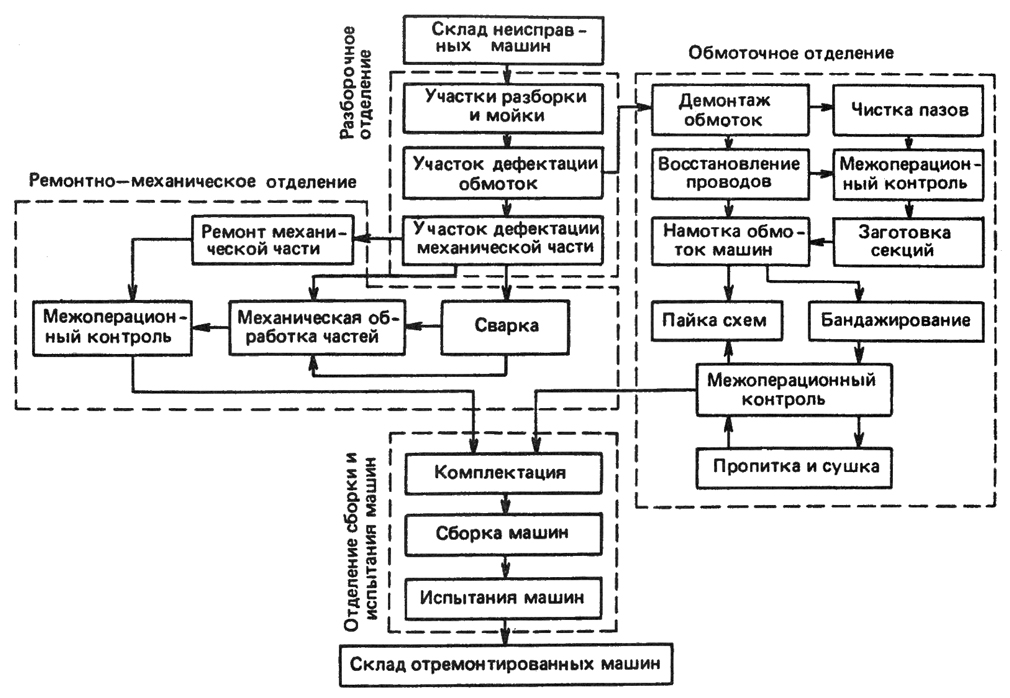 2.4).
2.4).
Исследования мутаций выявили ряд генов E. coli , которые при инактивации вызывают дефекты гомологичной рекомбинации, указывая на то, что их белковые продукты каким-то образом участвуют в этом процессе. Были описаны три различные системы рекомбинации, это пути RecBCD, RecE и RecF, причем RecBCD, по-видимому, является наиболее важным в бактериях (Camerini-Otero and Hsieh, 1995). В этом пути рекомбинация инициируется ферментом RecBCD, который обладает как нуклеазной, так и геликазной активностью.Его точный механизм действия неизвестен: в простейшей модели фермент связывается с одним концом линейной молекулы и раскручивает ее, пока не достигнет первой копии консенсусной последовательности из восьми нуклеотидов 5′-GCTGGTGG-3′ (которая довольно путано называется chi-сайт), который встречается один раз каждые 6 т.п.н. в ДНК E. coli (Blattner et al. ., 1997). Затем нуклеазная активность фермента создает одноцепочечный разрыв в положении примерно 56 нуклеотидов к 3′-стороне хи-сайта (). В альтернативных предложениях фермент RecBCD создает разрывы по мере продвижения вдоль линейной ДНК, причем эта активность ингибируется при достижении сайта chi, причем последний из этих прогрессирующих разрывов эквивалентен одиночному разрыву, предусмотренному в первой модели (Eggleston and West, 1996). ).
В альтернативных предложениях фермент RecBCD создает разрывы по мере продвижения вдоль линейной ДНК, причем эта активность ингибируется при достижении сайта chi, причем последний из этих прогрессирующих разрывов эквивалентен одиночному разрыву, предусмотренному в первой модели (Eggleston and West, 1996). ).
Рисунок 14.30
Путь RecBCD для гомологичной рекомбинации в Escherichia coli . Показаны события, приводящие к образованию гетеродуплекса. В нижней структуре нить ДНК, покрытая RecA, сформировала тройную структуру, которая считается промежуточной (более подробно)…)
Каким бы ни был точный механизм, фермент RecBCD производит свободный одноцепочечный конец, который, в соответствии с модификацией Мезельсона-Раддинга, вторгается в интактный партнер, в данном случае в кольцевой геном E. coli . Эта стадия опосредована белком RecA, который образует покрытую белком нить ДНК, способную внедряться в интактную двойную спираль и образовывать D-петлю (см. ). Промежуточным звеном в образовании D-петли, вероятно, является тройная структура, трехцепочечная спираль ДНК, в которой вторгшийся полинуклеотид лежит в пределах большой бороздки интактной спирали и образует водородные связи с парами оснований, с которыми он сталкивается (Camerini-Otero и Се, 1995).
). Промежуточным звеном в образовании D-петли, вероятно, является тройная структура, трехцепочечная спираль ДНК, в которой вторгшийся полинуклеотид лежит в пределах большой бороздки интактной спирали и образует водородные связи с парами оснований, с которыми он сталкивается (Camerini-Otero и Се, 1995).
Миграция ветвей катализируется белками RuvA и RuvB, оба из которых прикрепляются к точке ветвления структуры Холлидея. Рентгеноструктурные исследования предполагают, что четыре копии RuvA связываются непосредственно с ветвью, образуя ядро, к которому прикрепляются два кольца RuvB, каждое из которых состоит из восьми белков, по одному с каждой стороны (; Rafferty et al. ., 1996). Получившаяся структура могла бы действовать как «молекулярный двигатель», вращая спирали нужным образом, чтобы точка ветвления двигалась.Белок RecG также играет роль в миграции ветвей, но неясно, связано ли это с RuvAB или как часть альтернативного механизма (Eggleston and West, 1996).
Рисунок 14.
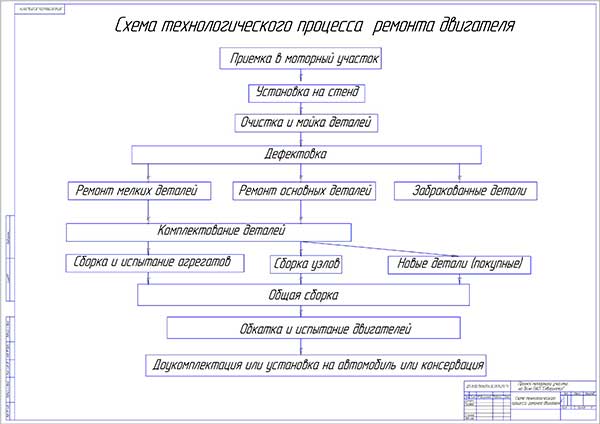 31
31Роль белков Ruv в гомологичной рекомбинации у Escherichia coli. Миграция ветвей индуцируется структурой, состоящей из четырех копий RuvA, связанных с соединением Холлидея с кольцом RuvB с каждой стороны. После отсоединения RuvAB два RuvC (далее…)
Миграция ветки не выглядит случайным процессом, а вместо этого предпочтительно останавливается на последовательности
.Эта последовательность часто встречается в геноме E. coli , поэтому, по-видимому, миграция не останавливается при первом же достижении мотива. Когда миграция ветвей заканчивается, комплекс RuvAB отсоединяется и заменяется двумя белками RuvC (см. ), которые осуществляют расщепление, разрешающее структуру Холлидея. Разрезы делаются между вторым Т и компонентами последовательности распознавания.Модель двухцепочечного разрыва для рекомбинации у дрожжей
Хотя модель Холлидея для гомологичной рекомбинации, либо в ее исходной форме, либо в модификации Мезельсона и Раддинга, объясняет большинство результатов рекомбинации во всех организмах, она имеет несколько недостатков , что побудило к разработке альтернативных схем.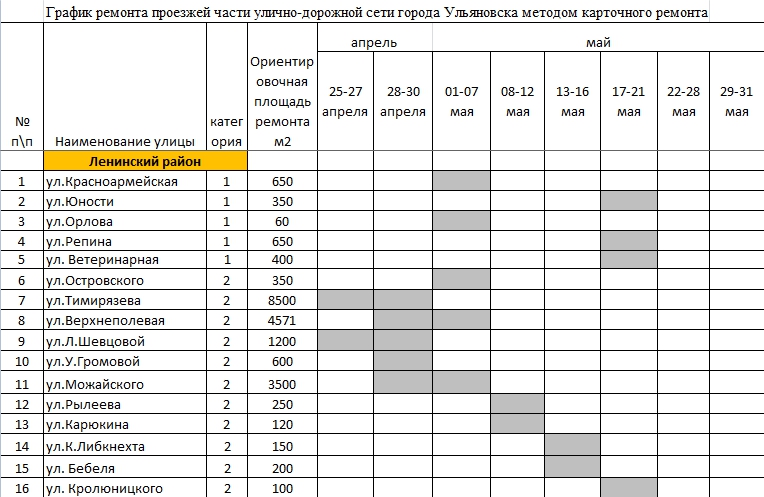 В частности, считалось, что модель Холлидея не может объяснить конверсию генов — явление, впервые описанное у дрожжей и грибов, но теперь известное для многих эукариот. У дрожжей слияние пары гамет приводит к образованию зиготы, которая дает начало аску, содержащему четыре гаплоидные споры, генотипы которых можно определить индивидуально. Если гаметы имеют разные аллели в определенном локусе, то при нормальных обстоятельствах две споры будут демонстрировать один генотип, а две — другой генотип, но иногда эта ожидаемая схема сегрегации 2 : 2 заменяется неожиданным соотношением 3 : 1. .Это называется генной конверсией, потому что такое соотношение можно объяснить только «конверсией» одного из аллелей из одного типа в другой, предположительно путем рекомбинации во время мейоза, который происходит после слияния гамет.
В частности, считалось, что модель Холлидея не может объяснить конверсию генов — явление, впервые описанное у дрожжей и грибов, но теперь известное для многих эукариот. У дрожжей слияние пары гамет приводит к образованию зиготы, которая дает начало аску, содержащему четыре гаплоидные споры, генотипы которых можно определить индивидуально. Если гаметы имеют разные аллели в определенном локусе, то при нормальных обстоятельствах две споры будут демонстрировать один генотип, а две — другой генотип, но иногда эта ожидаемая схема сегрегации 2 : 2 заменяется неожиданным соотношением 3 : 1. .Это называется генной конверсией, потому что такое соотношение можно объяснить только «конверсией» одного из аллелей из одного типа в другой, предположительно путем рекомбинации во время мейоза, который происходит после слияния гамет.
Рисунок 14.32
Конверсия генов. Одна гамета содержит аллель A , а другая содержит аллель a . Они сливаются, образуя зиготу, которая дает начало четырем гаплоидным спорам, содержащимся в одной аске. В норме две споры будут иметь аллель A , а две — аллель (more…)
В норме две споры будут иметь аллель A , а две — аллель (more…)
Модель двухцепочечного разрыва дает возможность генной конверсии происходить в процессе рекомбинации. Она начинается не с одноцепочечного разрыва, как в схеме Холлидея, а с двухцепочечного разреза, разрывающего одного из партнеров рекомбинации на две части (). Это может показаться радикальным шагом, но было показано, что белком, ответственным за разрез, является ДНК-топоизомераза типа II (раздел 13.1.2), которая образует ковалентные связи с двумя фрагментами ДНК и, следовательно, предотвращает их полный дрейф. отдельно.После двухцепочечного разреза одна цепь в каждой половине молекулы обрезается экзонуклеазой 5’→3′, поэтому каждый конец теперь имеет 3′-выступ примерно из 500 нуклеотидов. Один из них вторгается в гомологичную молекулу ДНК способом, подобным тому, который предусмотрен схемой Мезельсона-Раддинга, создавая соединение Холлидея, которое может мигрировать вдоль гетеродуплекса, если вторгающаяся цепь удлиняется ДНК-полимеразой. Для завершения гетеродуплекса также удлиняется другая оборванная нить (не участвующая в соединении Холлидея).Обратите внимание, что оба синтеза ДНК включают удлинение цепей от партнера, подвергшегося двухцепочечному разрезу, с использованием в качестве матриц эквивалентных областей неразрезанного партнера. Это лежит в основе генной конверсии, потому что это означает, что полинуклеотидные сегменты, удаленные экзонуклеазой от разрезанного партнера, были заменены копиями ДНК от неразрезанного партнера.
Для завершения гетеродуплекса также удлиняется другая оборванная нить (не участвующая в соединении Холлидея).Обратите внимание, что оба синтеза ДНК включают удлинение цепей от партнера, подвергшегося двухцепочечному разрезу, с использованием в качестве матриц эквивалентных областей неразрезанного партнера. Это лежит в основе генной конверсии, потому что это означает, что полинуклеотидные сегменты, удаленные экзонуклеазой от разрезанного партнера, были заменены копиями ДНК от неразрезанного партнера.
Рисунок 14.33
Модель двухцепочечного разрыва для рекомбинации в дрожжах. Эта модель объясняет, как может происходить конверсия генов.См текст для деталей.
Полученный гетеродуплекс имеет пару структур Холлидея, которые могут быть разрешены несколькими способами, некоторые из которых приводят к генной конверсии, а другие дают стандартный реципрокный обмен цепями. Пример, приводящий к конверсии генов, показан на рис.
Модель двухцепочечного разрыва была достаточно хорошо охарактеризована у дрожжей, чтобы можно было не сомневаться в том, что она происходит, по крайней мере, в форме, близкой к той, что показана на рис. Некоторые белки, участвующие в рекомбинации у дрожжей, очень похожи на свои аналоги в E.coli — эукариотический RAD51, например, имеет сходство последовательности с RecA и, как полагают, работает таким же образом (Baumann and West, 1998), что побуждает предположить, что рекомбинация во всех организмах происходит по системе двухцепочечного разрыва. Пока что имеется мало доказательств в поддержку этой идеи, особенно для более крупных хромосом высших эукариот, и многие генетики сопротивляются предположению, что ДНК позвоночных подвергается частым двухцепочечным разрывам во время мейоза.
Некоторые белки, участвующие в рекомбинации у дрожжей, очень похожи на свои аналоги в E.coli — эукариотический RAD51, например, имеет сходство последовательности с RecA и, как полагают, работает таким же образом (Baumann and West, 1998), что побуждает предположить, что рекомбинация во всех организмах происходит по системе двухцепочечного разрыва. Пока что имеется мало доказательств в поддержку этой идеи, особенно для более крупных хромосом высших эукариот, и многие генетики сопротивляются предположению, что ДНК позвоночных подвергается частым двухцепочечным разрывам во время мейоза.
Коробка 14.2
Пути рекомбинации RecE и RecF Escherichia coli . Пути RecBCD, RecE и RecF включают сходные механизмы и используют несколько одинаковых белков. RecA участвует в каждом пути, RecF является компонентом путей RecE и RecF, (подробнее…)
14.3.2. Сайт-специфическая рекомбинация
Наличие области обширной гомологии не является обязательным условием для рекомбинации: процесс может также инициироваться между двумя молекулами ДНК, которые имеют только очень короткие общие последовательности. Это называется сайт-специфической рекомбинацией, и она широко изучалась из-за роли, которую она играет во время инфекционного цикла бактериофага λ.
Это называется сайт-специфической рекомбинацией, и она широко изучалась из-за роли, которую она играет во время инфекционного цикла бактериофага λ.
Интеграция ДНК λ в геном
E. coli включает сайт-специфическую рекомбинацию. Один из них, литический путь, приводит к быстрому синтезу белков оболочки λ в сочетании с репликацией генома λ, что приводит к гибели бактерии и высвобождению новых фагов в течение примерно 20 минут после первоначального заражения.Напротив, если фаг следует лизогенному пути, новые фаги не появляются сразу. Бактерия делится как обычно, возможно, для многих клеточных делений, с фагом в покоящейся форме, называемой профагом. В конце концов, возможно, в результате повреждения ДНК или какого-либо другого стимула, фаг снова становится активным, реплицируя свой геном, направляя синтез белков оболочки и вырываясь из клетки. Во время лизогенной фазы геном λ интегрируется в геном E.coli хромосома. Поэтому он реплицируется всякий раз, когда копируется ДНК E.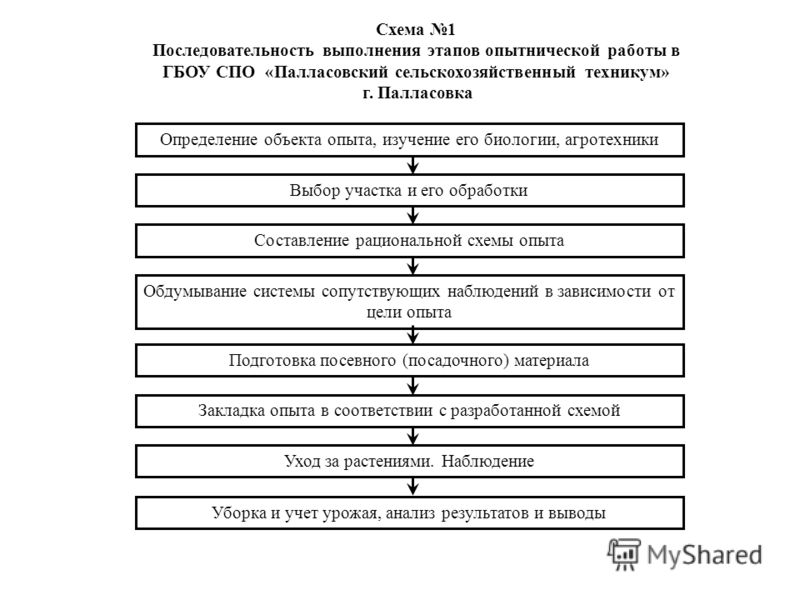 coli , и таким образом передается дочерним клеткам, как если бы это была стандартная часть генома бактерии. Интеграция происходит путем сайт-специфической рекомбинации между сайтами и , одним на λ-геноме и одним на хромосоме E. coli , которые имеют в своем центре идентичную последовательность из 15 п.н. Поскольку это рекомбинация между двумя кольцевыми молекулами, в результате образуется один больший круг; другими словами, ДНК λ интегрируется в бактериальный геном.Вторая сайт-специфическая рекомбинация между двумя сайтами и , теперь оба содержащимися в одной молекуле, обращает исходный процесс и высвобождает λ-ДНК, которая теперь может вернуться к литическому способу заражения и прямому синтезу новых фагов.
coli , и таким образом передается дочерним клеткам, как если бы это была стандартная часть генома бактерии. Интеграция происходит путем сайт-специфической рекомбинации между сайтами и , одним на λ-геноме и одним на хромосоме E. coli , которые имеют в своем центре идентичную последовательность из 15 п.н. Поскольку это рекомбинация между двумя кольцевыми молекулами, в результате образуется один больший круг; другими словами, ДНК λ интегрируется в бактериальный геном.Вторая сайт-специфическая рекомбинация между двумя сайтами и , теперь оба содержащимися в одной молекуле, обращает исходный процесс и высвобождает λ-ДНК, которая теперь может вернуться к литическому способу заражения и прямому синтезу новых фагов.
Рисунок 14.34
Интеграция генома бактериофага λ в хромосомную ДНК Escherichia coli . И λ, и E. ДНК coli имеет копию сайта att , каждый из которых содержит идентичную центральную последовательность, называемую «O», и фланкирующие последовательности P (подробнее. ..)
..)
Рекомбинация катализируется специализированной топоизомеразой I типа (раздел 13.1.2), называемой интегразой (Kwon et al ., 1997), членом разнообразного семейства рекомбиназ, присутствующих в бактериях, археях и дрожжах. . Фермент делает двухцепочечный разрез в шахматном порядке в эквивалентных положениях в λ- и бактериальных и сайтах. Затем два коротких одноцепочечных выступа обмениваются между молекулами ДНК, образуя соединение Холлидея, которое перемещает несколько пар оснований вдоль гетеродуплекса перед расщеплением.Это расщепление, при условии, что оно выполнено в соответствующей ориентации, разрешает структуру Холлидея таким образом, что λ-ДНК встраивается в геном E. coli . Аналогичный процесс лежит в основе эксцизии, которая также осуществляется интегразой, но в сочетании со вторым белком, «эксцизионазой», кодируемым геном λ xis . Если бы интеграза могла выполнять вырезание сама по себе, то она, вероятно, вырезала бы λ-ДНК, как только интегрировала ее.
14.3.3. Транспозиция
Транспозиция — это не тип рекомбинации, а процесс, использующий рекомбинацию, конечным результатом которого является перенос сегмента ДНК из одного положения генома в другое. Характерной чертой транспозиции является то, что к переносимому сегменту примыкает пара коротких прямых повторов (), которые, как мы увидим, образуются в процессе транспозиции.
Рисунок 14.35
Интегрированные мобильные элементы окружены короткими прямыми повторяющимися последовательностями.Этот конкретный транспозон окружен тетрануклеотидным повтором 5′-CTGG-3′. Другие транспозоны имеют другую последовательность прямых повторов.
В разделе 2.4.2 мы рассмотрели различные типы мобильных элементов, известных у эукариот и прокариот, и обнаружили, что их можно разделить на три категории на основе их механизма транспозиции ():
- Транспозоны ДНК, которые транспонируются репликативно, при этом исходный транспозон остается на месте, а новая копия появляется в другом месте генома;
- Транспозоны ДНК, которые транспонируются консервативно, при этом исходный транспозон перемещается в новое место путем вырезания и вставки;
- Ретроэлементы, все из которых транспонируются через промежуточную РНК.

Рисунок 14.36
Репликативная и консервативная транспозиция. Транспозоны ДНК используют либо репликативный, либо консервативный путь (некоторые могут использовать оба). Ретроэлементы транспонируются репликативно через промежуточную РНК.
Теперь мы рассмотрим события рекомбинации, ответственные за каждый из этих трех типов транспозиции.
Графа 14.3
Метилирование и транспозиция ДНК. Транспозиция может иметь пагубные последствия для генома. Эти эффекты выходят за рамки очевидного нарушения активности гена, которое произойдет, если мобильный элемент займет новое положение, лежащее в кодирующей области (подробнее…)
Репликативная и консервативная транспозиция ДНК-транспозонов
За прошедшие годы был предложен ряд моделей репликативной и консервативной транспозиции, но большинство из них представляют собой модификации схемы, первоначально описанной Шапиро (1979). Согласно этой модели, репликативная транспозиция бактериального элемента, такого как транспозон типа Tn3 или мобильный фаг (раздел 2. 4.2), инициируется одной или несколькими эндонуклеазами, которые делают одноцепочечные разрезы по обе стороны от транспозона и в целевой сайт, куда будет вставлена новая копия элемента ().В месте-мишени два разреза разделены несколькими парами оснований, так что расщепленная двухцепочечная молекула имеет короткие выступающие 5′-концы.
4.2), инициируется одной или несколькими эндонуклеазами, которые делают одноцепочечные разрезы по обе стороны от транспозона и в целевой сайт, куда будет вставлена новая копия элемента ().В месте-мишени два разреза разделены несколькими парами оснований, так что расщепленная двухцепочечная молекула имеет короткие выступающие 5′-концы.
Рисунок 14.37
Модель процесса, приводящего к репликативной и консервативной транспозиции. См текст для деталей.
Лигирование этих 5′-выступов со свободными 3′-концами по обе стороны от транспозона дает гибридную молекулу, в которой исходные две ДНК — та, что содержит транспозон, и та, которая содержит сайт-мишень — связаны друг с другом мобильным элементом в окружении пары структур, напоминающих вилки репликации.Синтез ДНК на этих вилках репликации копирует мобильный элемент и превращает первоначальный гибрид в коинтеграт, в котором две исходные ДНК все еще связаны. Гомологическая рекомбинация между двумя копиями транспозона разъединяет коинтеграт, отделяя исходную молекулу ДНК (с ее копией транспозона, все еще на месте) от молекулы-мишени, которая теперь содержит копию транспозона. Таким образом, произошла репликативная транспозиция.
Таким образом, произошла репликативная транспозиция.
Модификация только что описанного процесса изменяет способ транспозиции с репликативного на консервативный (см. ).Вместо синтеза ДНК гибридная структура преобразуется обратно в две отдельные молекулы ДНК, просто создавая дополнительные одноцепочечные разрывы по обе стороны от транспозона. Это вырезает транспозон из его исходной молекулы, оставляя его «вставленным» в ДНК-мишень.
Транспозиция ретроэлементов
С точки зрения человека наиболее важными ретроэлементами являются ретровирусы, которые включают вирусы иммунодефицита человека, вызывающие СПИД, и различные другие вирулентные типы.Большая часть того, что мы знаем о ретротранспозиции, относится именно к ретровирусам, хотя считается, что другие ретротранспозоны, такие как ретротранспозоны семейств Ty1 / copya и Ty3 / gypsy , транспонируются по сходным механизмам.
Первым этапом ретротранспозиции является синтез РНК-копии вставленного ретроэлемента (). Длинный концевой повтор (LTR) на 5′-конце элемента содержит последовательность ТАТА, которая действует как промотор для транскрипции РНК-полимеразой II (раздел 9.2.2). Некоторые ретроэлементы также имеют энхансерные последовательности (раздел 9.3), которые, как считается, регулируют количество происходящей транскрипции. Транскрипция продолжается по всей длине элемента, вплоть до последовательности полиаденилирования (раздел 10.1.2) в 3′-LTR. Транскрипт теперь действует как матрица для РНК-зависимого синтеза ДНК, катализируемого ферментом обратной транскриптазы, кодируемым частью гена pol ретроэлемента (см. ). Поскольку это синтез ДНК, требуется праймер (раздел 13.2.2), и, как и при репликации генома, праймер состоит из РНК, а не из ДНК. Во время репликации генома de novo праймер синтезируется ферментом полимеразой (см. ), но ретроэлементы не кодируют РНК-полимеразы и поэтому не могут создавать праймеры таким образом. Вместо этого они используют одну из молекул тРНК клетки в качестве праймера, которая зависит от ретроэлемента: семейство элементов Ty1 / copy всегда использует тРНК Met , но другие ретроэлементы используют другие тРНК.
Длинный концевой повтор (LTR) на 5′-конце элемента содержит последовательность ТАТА, которая действует как промотор для транскрипции РНК-полимеразой II (раздел 9.2.2). Некоторые ретроэлементы также имеют энхансерные последовательности (раздел 9.3), которые, как считается, регулируют количество происходящей транскрипции. Транскрипция продолжается по всей длине элемента, вплоть до последовательности полиаденилирования (раздел 10.1.2) в 3′-LTR. Транскрипт теперь действует как матрица для РНК-зависимого синтеза ДНК, катализируемого ферментом обратной транскриптазы, кодируемым частью гена pol ретроэлемента (см. ). Поскольку это синтез ДНК, требуется праймер (раздел 13.2.2), и, как и при репликации генома, праймер состоит из РНК, а не из ДНК. Во время репликации генома de novo праймер синтезируется ферментом полимеразой (см. ), но ретроэлементы не кодируют РНК-полимеразы и поэтому не могут создавать праймеры таким образом. Вместо этого они используют одну из молекул тРНК клетки в качестве праймера, которая зависит от ретроэлемента: семейство элементов Ty1 / copy всегда использует тРНК Met , но другие ретроэлементы используют другие тРНК.
Рисунок 14.38
Транспозиция ретроэлемента – часть 1. На этой диаграмме показано, как интегрированный ретроэлемент копируется в свободную двухцепочечную версию ДНК. Первым этапом является синтез копии РНК, которая затем преобразуется в двухцепочечную ДНК с помощью ряда событий (подробнее…)
Праймер тРНК отжигается в сайте в пределах 5′ LTR (см. ) На первый взгляд это кажется странным местом для сайта праймирования, потому что это означает, что синтез ДНК направлен в сторону от центральной области ретроэлемента и, таким образом, приводит только к короткой копии части 5′ LTR.На самом деле, когда копия ДНК удлиняется до конца LTR, часть матрицы РНК деградирует, и образовавшийся выступ ДНК повторно отжигается до 3′-LTR ретроэлемента, который, будучи длинным концом повтор имеет ту же последовательность, что и 5′ LTR, и поэтому может образовывать пару оснований с копией ДНК. Синтез ДНК теперь продолжается вдоль матрицы РНК, в конечном итоге вытесняя праймер тРНК. Обратите внимание, что результатом является ДНК-копия всего шаблона, включая сайт праймирования: переключение шаблона, по сути, является стратегией, которую ретроэлемент использует для решения проблемы «укорочения концов», той же проблемы, которую хромосомные ДНК решают посредством синтез теломер (раздел 13.2.4).
Обратите внимание, что результатом является ДНК-копия всего шаблона, включая сайт праймирования: переключение шаблона, по сути, является стратегией, которую ретроэлемент использует для решения проблемы «укорочения концов», той же проблемы, которую хромосомные ДНК решают посредством синтез теломер (раздел 13.2.4).
Завершение синтеза первой нити ДНК приводит к гибриду ДНК-РНК. РНК частично расщепляется ферментом РНКазой H, кодируемым другой частью гена pol . Нерасщепленная РНК, обычно всего лишь один фрагмент, присоединенный к короткой полипуриновой последовательности, примыкающей к 3′-концу LTR, инициирует синтез второй цепи ДНК, опять же с помощью обратной транскриптазы, которая способна действовать как РНК-, так и ДНК-транскриптаза. -зависимая ДНК-полимераза. Как и в первом раунде синтеза ДНК, синтез второй цепи первоначально приводит к копии ДНК только LTR, но второй переключатель матрицы на другом конце молекулы позволяет удлинить копию ДНК до тех пор, пока она не станет полной. .Это создает матрицу для дальнейшего удлинения первой цепи ДНК, так что полученная двухцепочечная ДНК представляет собой полную копию внутренней области ретроэлемента плюс два LTR.
.Это создает матрицу для дальнейшего удлинения первой цепи ДНК, так что полученная двухцепочечная ДНК представляет собой полную копию внутренней области ретроэлемента плюс два LTR.
Осталось только вставить новую копию ретроэлемента в геном. Первоначально считалось, что вставка происходит случайным образом, но теперь выяснилось, что, хотя в качестве сайта-мишени не используется какая-либо конкретная последовательность, интеграция происходит предпочтительно в определенных положениях (Devine and Boeke, 1996).Вставка включает удаление двух нуклеотидов с 3′-концов двухцепочечного ретроэлемента ферментом интегразой (кодируется еще одной частью pol ). Интеграза также делает ступенчатые разрезы в геномной ДНК, так что и ретроэлемент, и сайт интеграции теперь имеют 5′-выступы (10). Эти выступы могут не иметь комплементарных последовательностей, но они все же каким-то образом взаимодействуют, так что ретроэлемент вставляется в геномную ДНК. Взаимодействие приводит к потере выступов ретроэлемента и заполнению оставшихся пробелов, что означает, что сайт интеграции дублируется в пару прямых повторов, по одному на каждом конце вставленного ретроэлемента.
Рисунок 14.39
Транспозиция ретроэлемента — Часть 2. Интеграция ретроэлемента в геном приводит к прямому повтору из четырех нуклеотидов по обе стороны от вставленной последовательности. Для ретровирусов на этом этапе пути транспозиции требуется как интеграза (подробнее…)
Типы, категории, варианты, профессии и контрольные списки работ по техническому обслуживанию — Управление цепочками поставок | Динамика 365
- Статья
- 19 минут на чтение
Пожалуйста, оцените свой опыт
да Нет
Любая дополнительная обратная связь?
Отзыв будет отправлен в Microsoft: при нажатии кнопки отправки ваш отзыв будет использован для улучшения продуктов и услуг Microsoft. Политика конфиденциальности.
Политика конфиденциальности.
Представлять на рассмотрение
Спасибо.
В этой статье
Тип актива привязан к каждому активу. Типы активов определяют типы работ по обслуживанию (и, следовательно, работы по обслуживанию), которые могут выполняться на активах. При создании рабочего задания необходимо выбрать тип задания обслуживания. Можно выбрать только те типы заданий обслуживания, которые связаны с настройкой типа актива, используемого для актива.
Графический обзор активов и типов заданий по обслуживанию, а также их связь с рабочими заданиями см. в разделе Функциональные местоположения и активы.
Варианты типа задания обслуживания можно настроить для типа задания обслуживания. Варианты типа задания обслуживания определяют варианты типа задания, такие как размеры (маленький, средний или большой), периоды (еженедельно, раз в две недели, один месяц или три месяца) и конфигурации (низкостандартная, гибкая или высокопроизводительная).
Профессии по техническому обслуживанию предоставляют информацию о профессиональных профессиях, таких как механика, электрика и гидравлика.Требования к компетенциям могут быть установлены в отношении работы по техническому обслуживанию. Все виды работ по техническому обслуживанию могут использоваться в отношении всех типов работ по техническому обслуживанию. Выбор варианта типа работы по техническому обслуживанию и/или обмена работами по техническому обслуживанию в рабочем задании является необязательным.
Для каждого типа задания обслуживания можно создать варианты настройки типа задания обслуживания. Например, если у вас есть тип задания по техническому обслуживанию с именем Сервис , вы можете создать следующие варианты для этого типа задания по техническому обслуживанию: Грузовые автомобили 30 000 км , Легковые автомобили 30 000 км и Фургоны 30 000 км .
Категории типов работ по техобслуживанию используются для сбора группы типов работ по техобслуживанию в обзорных целях. Примеры категорий типов работ по техническому обслуживанию могут включать Калибровка , Осмотр , Капитальный ремонт и Контрольно-измерительные приборы .
Примеры категорий типов работ по техническому обслуживанию могут включать Калибровка , Осмотр , Капитальный ремонт и Контрольно-измерительные приборы .
Шаблоны контрольных списков обслуживания и переменные контрольных списков обслуживания используются для настройки контрольных списков обслуживания. Контрольные списки обслуживания настраиваются для типов работ по обслуживанию и используются в рабочих заданиях.
Сначала вы устанавливаете требуемые категории типов работ по техническому обслуживанию, варианты типов работ по техническому обслуживанию и обмены работами по техническому обслуживанию. Затем вы создаете типы заданий обслуживания. Наконец, на странице Тип задания обслуживания по умолчанию вы создаете все варианты типов заданий обслуживания, которые требуются для вашего оборудования. На этой странице вы также можете настроить прогнозы, контрольные списки обслуживания и инструменты для комбинации типов работ по обслуживанию.
Примечание
Тип задания обслуживания может быть связан только с одной категорией типа задания обслуживания.
Создать категорию типа задания обслуживания
Выберите Управление активами > Настройка > Задания > Категории типов заданий обслуживания .
Выберите Новый .
В поле Тип задания обслуживания введите идентификатор категории типа задания обслуживания.
В поле Имя введите имя.
После связывания категорий типов работ по обслуживанию с типами работ по обслуживанию в поле Типы работ отображается количество типов работ по обслуживанию, связанных с этой категорией типов работ по обслуживанию.
Создать вариант типа задания обслуживания
Выберите Управление активами > Настройка > Задания > Варианты типов заданий обслуживания .
Выберите Новый .
В поле Вариант типа задания обслуживания введите идентификатор варианта типа задания обслуживания.
В поле Описание введите описание.
На экспресс-вкладке Типы заданий обслуживания выберите Добавить , чтобы добавить тип задания обслуживания.
В поле Тип задания обслуживания выберите тип задания обслуживания.
Повторите шаги с 5 по 6, чтобы добавить дополнительные типы заданий обслуживания в вариант типа задания обслуживания.
На экспресс-вкладке Подробности поле Типы заданий показывает количество типов заданий обслуживания, которые были добавлены к этому варианту типа задания обслуживания.

Создать сделку по ремонту
Выберите Управление активами > Настройка > Работа > Торговля ремонтными работами .
Выберите Новый .
В поле Сделка введите идентификатор сделки по ремонту.
В поле Описание введите описание.
На экспресс-вкладке Навыки выберите Добавить , чтобы добавить новый навык, который должен быть связан с работой по техническому обслуживанию.
В поле Навык выберите навык.
В поле Уровень выберите уровень навыка.
Повторите шаги с 5 по 7, чтобы добавить больше навыков в работу по техническому обслуживанию.
На экспресс-вкладке Details поле Skills показывает количество навыков, которые были добавлены к этой профессии обслуживания.
На экспресс-вкладке Сертификаты выберите Добавить , чтобы добавить сертификат в сделку по обслуживанию.
В поле Тип сертификата выберите сертификат.
Повторите шаги с 9 по 10, чтобы добавить дополнительные сертификаты в обмен на обслуживание.
На экспресс-вкладке Подробности поле Сертификаты показывает количество сертификатов, которые были добавлены к этой сделке по обслуживанию.

Создать переменную контрольного списка обслуживания
При создании строк контрольного списка обслуживания в типе задания обслуживания по умолчанию необходимо выбрать тип контрольного списка обслуживания. Переменная — это один из типов контрольного списка технического обслуживания. Он используется для определения возможного результата в строке контрольного списка обслуживания, которая связана со строкой заказа на работу. Переменная позволяет вам создать набор предопределенных результатов без необходимости точного измерения.
Пример 1: Уровень масла можно измерить, задав три значения: Слишком высокий уровень , Слишком низкий уровень и Уровень в пределах диапазона . Для каждого значения вы определяете, будет ли результатом значения Pass , Fail или None .
Пример 2: Вы проводите визуальный осмотр оборудования для оценки износа.
Выберите Управление активами > Настройка > Контрольные списки обслуживания > Переменные контрольного списка обслуживания .
Выберите Новый .
В поле Переменная введите идентификатор переменной контрольного списка обслуживания.
В поле Имя введите имя.
На экспресс-вкладке Общие выберите Добавить , чтобы добавить строку для переменной.
Порядковый номер строки автоматически вводится в поле Номер строки . После того, как вы добавили все строки, вы можете изменить их номера по своему усмотрению.
Когда вы выбираете строку, а затем нажимаете клавишу со стрелкой вниз , следующий номер в последовательности автоматически вводится в следующей строке.В поле Значение введите описание значения.
В поле Результат выберите результат для строки.
 Когда вы выбираете строку, а затем нажимаете клавишу со стрелкой вниз , следующий номер в последовательности автоматически вводится в следующей строке.
Когда вы выбираете строку, а затем нажимаете клавишу со стрелкой вниз , следующий номер в последовательности автоматически вводится в следующей строке.Создайте шаблон контрольного списка обслуживания
Шаблоны контрольных списков технического обслуживания можно использовать в качестве общего набора задач, которые работник должен выполнить для правильного выполнения заказа на работу. Ссылка на шаблоны содержится в строках контрольного списка обслуживания в типе задания обслуживания по умолчанию. На шаблоны можно ссылаться в нескольких строках по умолчанию типа задания обслуживания.Таким образом, вы можете легко повторно использовать набор общих задач контрольного списка обслуживания. Примеры шаблонов контрольных списков технического обслуживания включают общие инструкции по технике безопасности, а также список элементов и условий, которые необходимо проверить на конкретном насосе или аналогичных моделях конвейерной ленты.
Выберите Управление активами > Настройка > Контрольные списки обслуживания > Шаблоны контрольных списков обслуживания .
Выберите Новый .
Идентификатор шаблона автоматически вводится в поле шаблона контрольного списка обслуживания .
В поле Имя введите имя шаблона контрольного списка обслуживания.
На экспресс-вкладке Строки контрольного списка обслуживания выберите Добавить , чтобы добавить строку шаблона.
Порядковый номер строки автоматически вводится в поле Номер строки . После того, как вы добавили все строки, вы можете изменить их номера по своему усмотрению.Когда вы выбираете строку, а затем нажимаете клавишу со стрелкой вниз , следующий номер в последовательности автоматически вводится в следующей строке.
В поле Тип выберите тип строки контрольного списка обслуживания. Для каждого типа контрольного списка обслуживания на экспресс-вкладке Сведения о строке отображаются связанные поля. Доступны следующие значения:
Текст — В строке есть текст, описывающий, что делать. Используйте этот тип контрольного списка обслуживания, если вы хотите, чтобы работник что-то проверил или осмотрел, но вы не ожидаете конкретного (измеримого) результата.После выбора этого типа введите имя или заголовок в поле Имя . В поле Инструкции введите описание того, что необходимо сделать. Если шаг является обязательным для контрольного списка обслуживания, установите для параметра Обязательный значение Да .
Заголовок — строка используется в качестве заголовка для группировки строк контрольного списка обслуживания, которые отображаются под ней.
Этот тип полезен, если у вас есть несколько строк контрольного списка обслуживания, которые можно разделить на определенные области.Заголовки предоставляют обзор для работника, который заполнит контрольный список обслуживания, содержащий много строк контрольного списка обслуживания. После выбора этого типа введите описательное имя в поле Имя .Шаблон – Строка используется для ссылки на существующий шаблон. После выбора этого типа введите имя шаблона в поле Имя . В поле Шаблон выберите шаблон.
Переменная – Строка используется для определения возможного результата в диапазоне.Сведения о настройке переменных контрольного списка обслуживания см. в разделе Создание переменной контрольного списка обслуживания. После выбора этого типа введите описательное имя переменной в поле Имя . В поле Переменная выберите переменную. В поле Инструкции введите описание того, что необходимо сделать.
Если шаг является обязательным для контрольного списка обслуживания, установите для параметра Обязательный значение Да .Измерение – Линия используется для записи определенного измерения.Вы можете настроить измерение, которое должно быть связано с предопределенным счетчиком. После выбора этого типа введите имя шаблона в поле Имя . Если этот шаг является обязательным для контрольного списка обслуживания, установите для параметра Обязательный значение Да . Если вы хотите использовать линию измерения в качестве регистрации счетчика, выберите счетчик в поле Счетчик . Затем соответствующее поле Unit автоматически обновляется. Если вы выбрали счетчик, выберите метод обновления в поле Value .В мин. значение и Макс. значение полей, введите допустимый диапазон значений. В поле Инструкции введите описание того, что необходимо сделать.
Примечание
Любая строка типа Измерение , не имеющая настройки счетчика, рассматривается как независимая регистрация измерения, для которой нет автоматического отслеживания в Управлении активами. Аналогичным образом, если выбранный тип счетчика отсутствует в активе, связанном с рабочим заданием, задача контрольного списка обслуживания рассматривается как независимое измерение.Значение счетчика может быть изменено несколько раз. Он не публикуется до тех пор, пока состояние жизненного цикла заказа на работу не будет изменено на состояние, в котором для параметра Контрольный список обслуживания процесса установлено значение Да .
На экспресс-вкладке Подробности поле Проверки показывает общее количество строк контрольного списка в вашем шаблоне. Это число включает вложенные строки в любом существующем шаблоне, на который вы ссылаетесь в своем шаблоне.
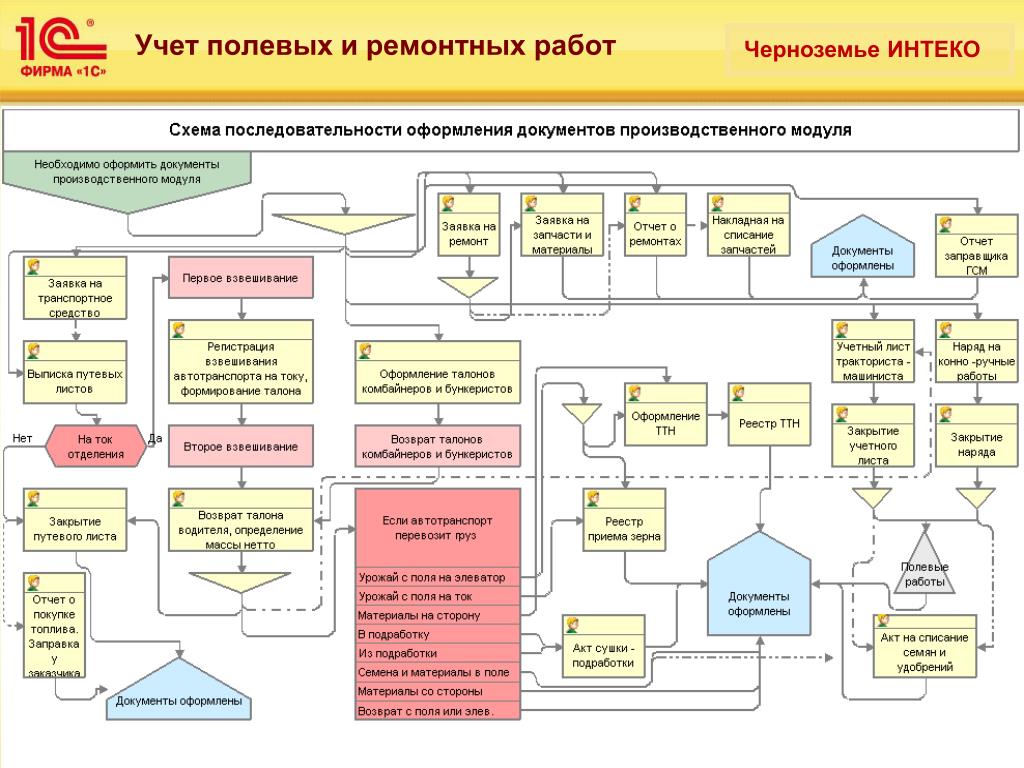
 Этот тип полезен, если у вас есть несколько строк контрольного списка обслуживания, которые можно разделить на определенные области.Заголовки предоставляют обзор для работника, который заполнит контрольный список обслуживания, содержащий много строк контрольного списка обслуживания. После выбора этого типа введите описательное имя в поле Имя .
Этот тип полезен, если у вас есть несколько строк контрольного списка обслуживания, которые можно разделить на определенные области.Заголовки предоставляют обзор для работника, который заполнит контрольный список обслуживания, содержащий много строк контрольного списка обслуживания. После выбора этого типа введите описательное имя в поле Имя . Если шаг является обязательным для контрольного списка обслуживания, установите для параметра Обязательный значение Да .
Если шаг является обязательным для контрольного списка обслуживания, установите для параметра Обязательный значение Да .
Создайте тип задания обслуживания
Выберите Управление активами > Настройка > Задания > Типы заданий обслуживания .
Выберите Новый .
В поле Тип задания обслуживания введите идентификатор типа задания обслуживания.
В поле Имя введите имя.
На экспресс-вкладке Details отображается обзор количества вариантов типов заданий обслуживания, навыков, сертификатов, последующих заданий и типов активов, созданных для этого типа заданий обслуживания. В поле Строки настройки отображается количество строк по умолчанию для типа задания обслуживания, которые были настроены для этого типа задания обслуживания.Поле Assets показывает количество активных активов, которые в настоящее время используют этот тип задания обслуживания.
На экспресс-вкладке Общие в поле Тип задания обслуживания выберите категорию типа задания обслуживания.
Установите для параметра Действия по простою обслуживания значение Да , если тип задания обслуживания требует остановки оборудования перед выполнением задания.
На экспресс-вкладке Описание введите описание типа задания обслуживания.
На экспресс-вкладке Варианты типов заданий обслуживания можно добавить варианты к типу заданий обслуживания.
На экспресс-вкладках Требуемые навыки и Требуемые сертификаты можно добавить требования к навыкам и сертификатам к типу работ по техническому обслуживанию.
Если следующим заданием по обслуживанию должен быть выполнен определенный тип, добавьте его на экспресс-вкладку Выполненные задания . Вы также можете настроить вариант типа задания на техническое обслуживание и сделку, связанную с типом задания на техническое обслуживание.Если последующее задание должно начаться за определенное количество дней до или после запуска задания, использующего этот тип задания обслуживания, введите количество дней в поле Задержка на дни .
Положительные числа представляют дни после начала связанного задания, а отрицательные числа представляют дни до запланированного начала связанного задания. Например, если ввести 5 , последующее задание начнется через пять дней после начала задания, относящегося к типу задания обслуживания.Если вы введете -3 , последующее задание начнется за три дня до запланированного запуска задания, относящегося к типу задания обслуживания.Примечание
Если вы добавите более одной строки типа задания обслуживания, последовательность строк указывает порядок, в котором они должны выполняться. Последовательность начинается в верхней части списка.
На экспресс-вкладке Типы активов можно добавить типы активов к типу задания обслуживания.


 Положительные числа представляют дни после начала связанного задания, а отрицательные числа представляют дни до запланированного начала связанного задания. Например, если ввести 5 , последующее задание начнется через пять дней после начала задания, относящегося к типу задания обслуживания.Если вы введете -3 , последующее задание начнется за три дня до запланированного запуска задания, относящегося к типу задания обслуживания.
Положительные числа представляют дни после начала связанного задания, а отрицательные числа представляют дни до запланированного начала связанного задания. Например, если ввести 5 , последующее задание начнется через пять дней после начала задания, относящегося к типу задания обслуживания.Если вы введете -3 , последующее задание начнется за три дня до запланированного запуска задания, относящегося к типу задания обслуживания.Создание строк по умолчанию для типов работ по обслуживанию и соответствующих прогнозов, контрольных списков обслуживания, инструментов, описаний и вложений
Выберите Управление активами > Настройка > Задания > Тип задания обслуживания по умолчанию .
–или–
Выберите Управление активами > Настройка > Задания > Типы заданий обслуживания , выберите тип задания обслуживания, а затем выберите Тип задания обслуживания по умолчанию .
Выберите Новый .
В полях Техническое место , Тип актива , Изготовитель , Модель и Актив выберите соответствующие значения в зависимости от того, насколько конкретным должен быть тип задания обслуживания по умолчанию.
В поле Тип задания обслуживания выберите тип задания обслуживания, если он не был выбран автоматически.
В полях Тип работы по техническому обслуживанию вариант и Торговля выберите нужный вариант типа работы по техническому обслуживанию и сделку по обслуживанию.
Выберите Прогноз .
На странице прогноза по умолчанию типа задания технического обслуживания можно делать прогнозы по часам, номенклатурам и расходам.На соответствующих вкладках выберите Добавить и сделайте выбор, чтобы создать необходимые прогнозы для типа задания обслуживания.
На вкладке Прогноз номенклатуры можно выбрать складские аналитики, которые должны отображаться в строке номенклатуры. Выберите Инвентаризация > Измерения , выберите размеры для отображения, установите для параметра Сохранить настройку значение Да , а затем выберите OK .
На вкладке Прогноз элемента выберите Элемент, где использовался , чтобы просмотреть обзор того, где элемент в выбранной строке используется в Управлении активами в отношении активов, типов работ по обслуживанию по умолчанию, запасных частей и рабочих заданий.
Экспресс-вкладка Итоги прогноза обслуживания показывает обзор итогов прогноза. Этот обзор включает общее количество часов и созданных строк прогноза.
Примечание
Чтобы скопировать настройку прогноза из другого типа задания обслуживания, выберите Копировать прогноз , а затем выберите тип задания обслуживания, из которого нужно скопировать настройку.
Выберите Сохранить , чтобы сохранить изменения.
Закройте страницу прогноза по умолчанию для типа задания обслуживания , чтобы вернуться на страницу по умолчанию для типа задания обслуживания .
Выберите Контрольный список технического обслуживания .
На странице Контрольный список по умолчанию для типа задания обслуживания можно добавить строки контрольного списка обслуживания к выбранному типу задания обслуживания по умолчанию.
Номера строк На экспресс-вкладке Строки проверки обслуживания выберите Новый , чтобы добавить строку контрольного списка обслуживания.автоматически вводятся в поле Номер строки , чтобы указать последовательность строк контрольного списка обслуживания.Вы можете редактировать номера строк по своему усмотрению. После создания первой строки контрольного списка обслуживания выберите строку, а затем нажмите клавишу со стрелкой вниз , чтобы добавить строку под ней. Кроме того, вы можете выбрать строку контрольного списка обслуживания, а затем выбрать New . В этом случае над выбранной строкой контрольного списка обслуживания добавляется новая строка.
В поле Тип выберите тип строки, а затем добавьте информацию, связанную с типом контрольного списка обслуживания.Описание доступных типов и связанных полей см. в разделе Создание шаблона контрольного списка обслуживания.
Примечание
Чтобы скопировать настройку контрольного списка обслуживания из другого типа задания обслуживания, выберите Копировать контрольный список обслуживания , а затем выберите тип задания обслуживания, из которого необходимо скопировать настройку.
Вы можете легко создать шаблон из существующего контрольного списка обслуживания. Затем вы можете повторно использовать шаблон в нескольких контрольных списках обслуживания. Новый шаблон будет точной копией действующего контрольного списка обслуживания.Выберите Создать шаблон и введите имя шаблона. Чтобы заменить существующий контрольный список обслуживания одной строкой со ссылкой на новый шаблон, установите для параметра Заменить значение Да . Содержимое шаблона можно просмотреть на странице сведений о шаблонах контрольных списков обслуживания .
Выберите Сохранить , чтобы сохранить изменения.
Вернуться на страницу типа задания обслуживания по умолчанию .
Выберите Инструменты .
На странице инструментов по умолчанию для типа задания обслуживания можно добавить инструменты (ресурсы), которые следует использовать для типа задания обслуживания. Выберите New , а затем выберите инструмент в поле Resource .
Примечание
Чтобы скопировать настройку инструмента из другого типа задания обслуживания, выберите Инструменты копирования , а затем выберите тип задания обслуживания, из которого нужно скопировать настройку.
Выберите Сохранить , чтобы сохранить изменения.
Вернуться на страницу типа задания обслуживания по умолчанию .
Выберите Описание работы .
На странице Описание работы выберите Изменить , а затем добавьте описание, связанное с выбранным типом задания обслуживания по умолчанию, как вам нужно.
Выберите Сохранить , чтобы сохранить описание.
Если вы добавите сюда описание работы, оно переопределит любое описание, настроенное для типа задания обслуживания на странице Типы заданий обслуживания . Если вы не добавите здесь описание работы, будет использоваться любое описание, настроенное для типа задания обслуживания. Описания автоматически переносятся в заказы на работу, в которых используется тип задания обслуживания или тип задания обслуживания по умолчанию.
Вернуться на страницу типа задания обслуживания по умолчанию .
Чтобы настроить вложения в строке по умолчанию для выбранного типа задания на обслуживание, выберите Прикрепить документы . Вложения, настроенные в строке по умолчанию для типа задания обслуживания, автоматически включаются в строки рабочих заданий, в которых используется строка по умолчанию для этого типа задания обслуживания.
Выберите Новый , а затем выберите тип документа.
Загрузите документ или файл.
Задайте поля на странице Приложения .Настройка вложения использует стандартные функции настройки документа.
Выберите Сохранить , чтобы сохранить вложение.
Примечание
Вложения в строке по умолчанию типа задания на обслуживание печатаются вместе с отчетом о рабочем задании, только если типы документов вложений выбраны на вкладке Типы документов на странице Параметры управления активами ( Управление активами > Настройка > Параметры управления активами ).Примеры вложений включают рекомендации, поясняющие, как выполнить определенное задание, или предопределенный контрольный список обслуживания (если вы не используете функцию контрольного списка обслуживания для строк по умолчанию типа задания обслуживания).
На странице Тип задания обслуживания по умолчанию каждая строка показывает количество прогнозируемых часов, а также количество строк, созданных для элементов, расходов, контрольных списков обслуживания и инструментов. Поле Assets показывает количество активных активов, связанных со строкой по умолчанию типа задания обслуживания.
Чтобы скопировать тип задания обслуживания по умолчанию в другой тип задания обслуживания по умолчанию, выберите строку по умолчанию типа задания обслуживания, чтобы скопировать другую настройку, выберите Копировать настройку , а затем выберите тип задания обслуживания по умолчанию для копирования.
Чтобы просмотреть список активов, планов обслуживания или циклов обслуживания, которые в настоящее время используют строку по умолчанию типа задания обслуживания, выберите строку, а затем выберите Используется .

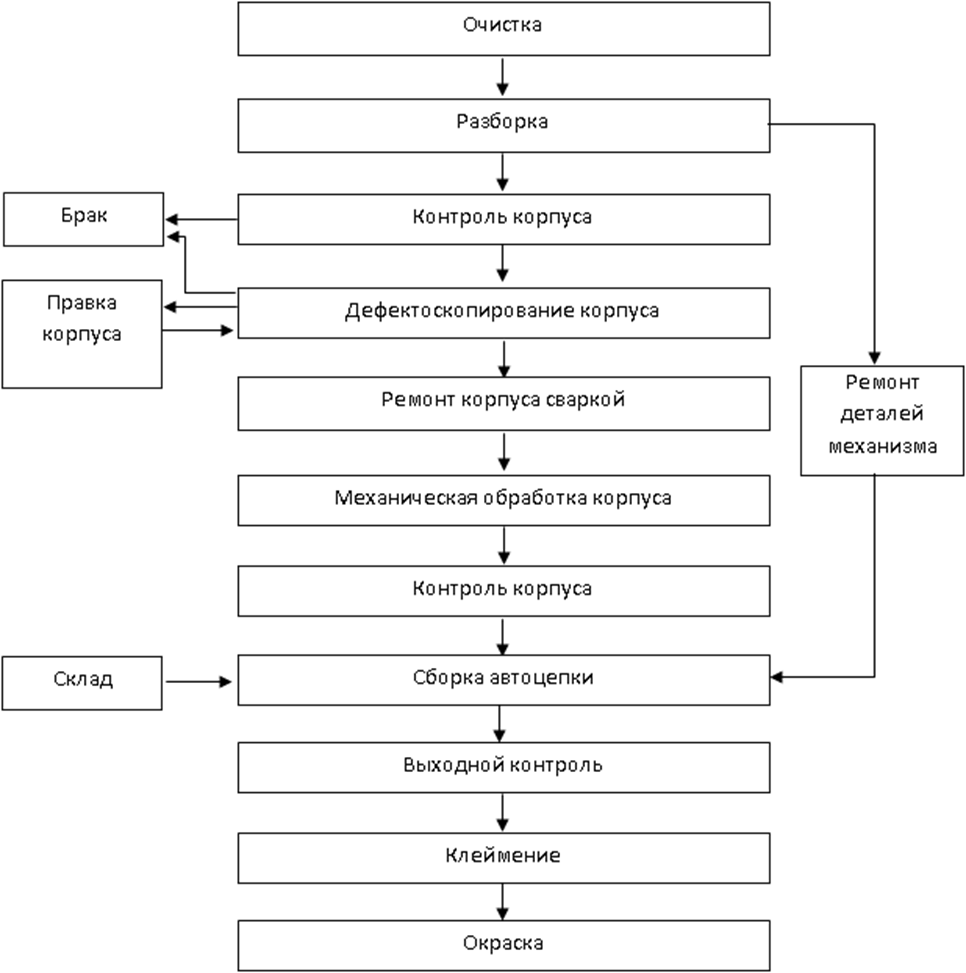

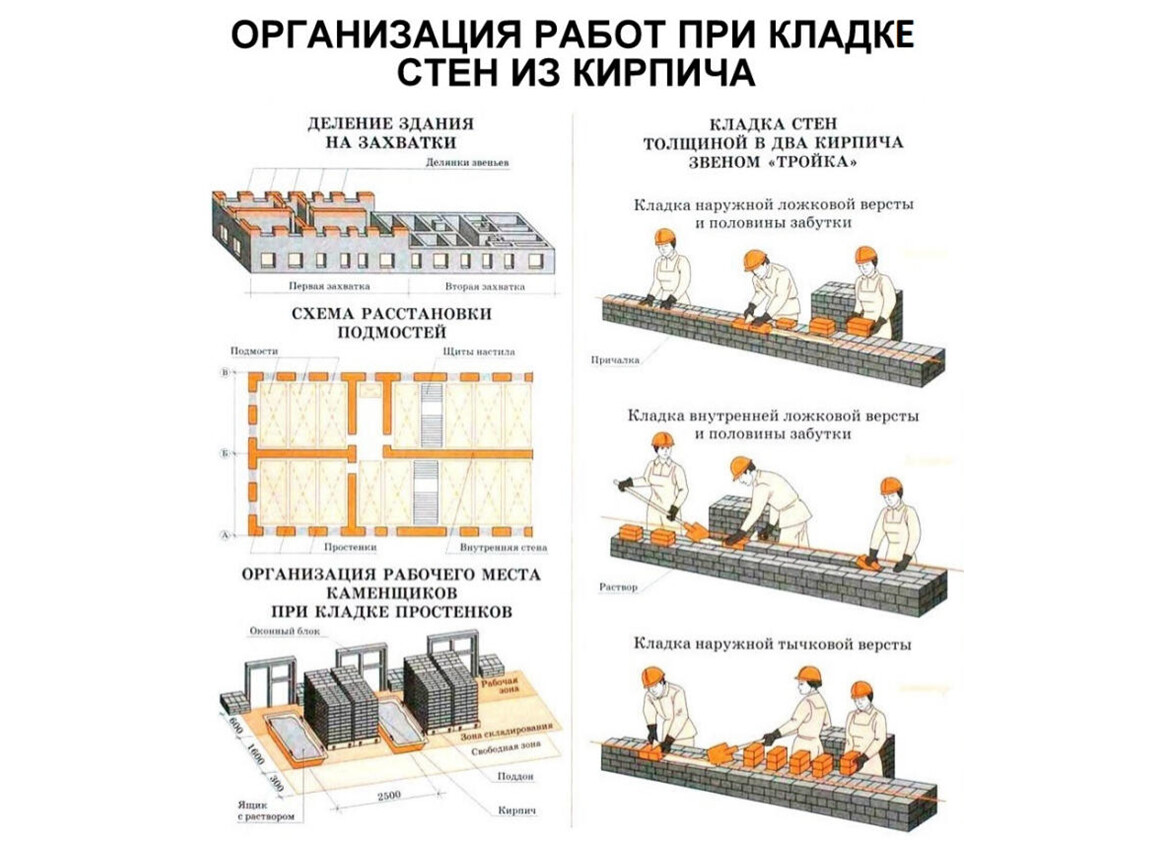 На экспресс-вкладке Строки проверки обслуживания выберите Новый , чтобы добавить строку контрольного списка обслуживания.
На экспресс-вкладке Строки проверки обслуживания выберите Новый , чтобы добавить строку контрольного списка обслуживания.
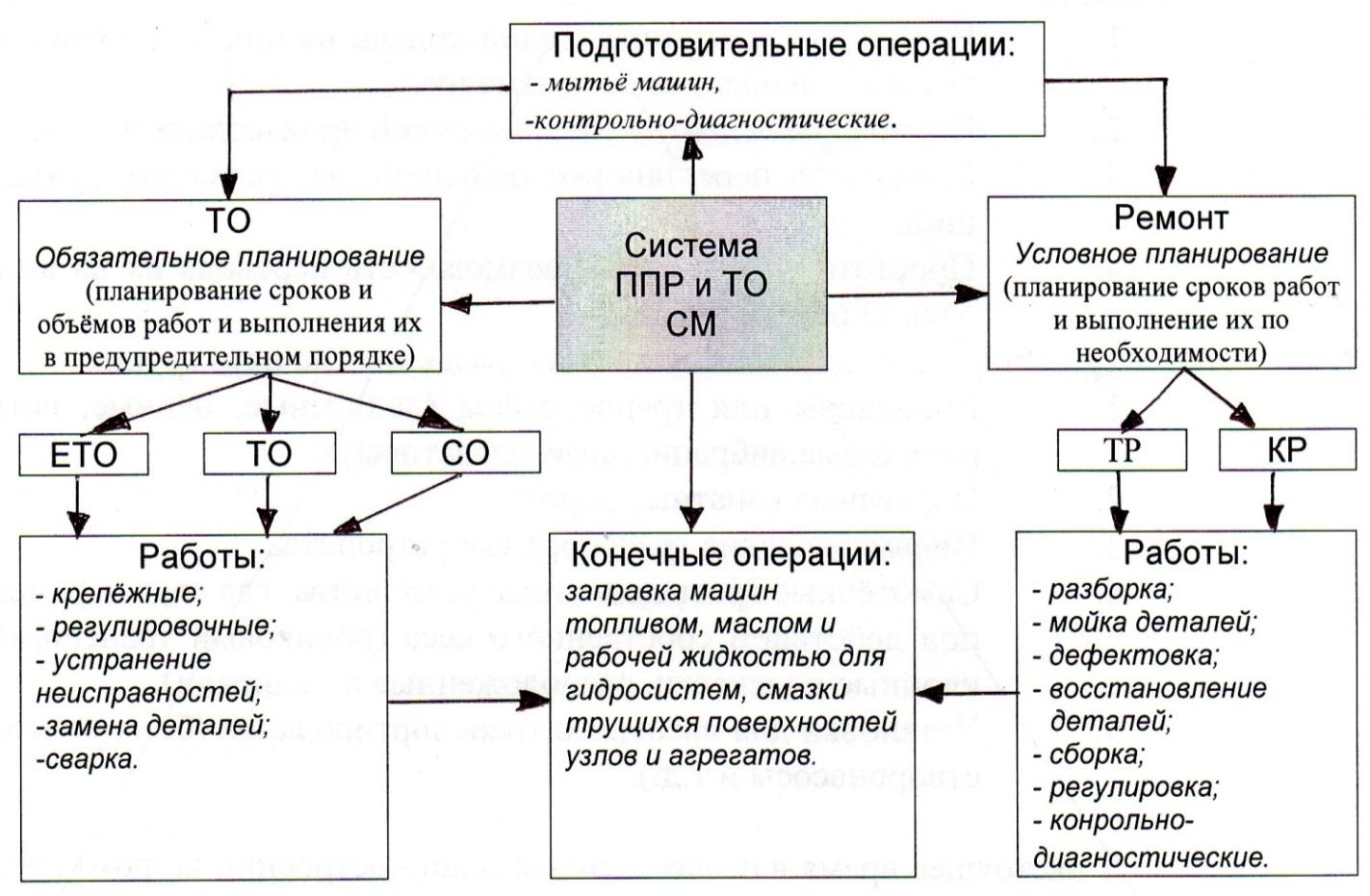

Когда система выбирает доступный тип задания по обслуживанию по умолчанию, который следует использовать в строке заказа на работу, выбор основывается на активе и настройке связанного типа актива.Управление активами просматривает все записи по умолчанию для типов работ по обслуживанию, которые связаны с типом работ по обслуживанию, связанным с типом активов, чтобы проверить возможное совпадение. Он всегда сначала проверяет наиболее конкретную комбинацию. Другими словами, чтобы найти наиболее конкретную комбинацию, Управление активами сначала проверяет возможное совпадение для поля Trade . Если совпадений не найдено, проверяется совпадение для поля варианта задания типа обслуживания . Если совпадений не найдено, он проверяет соответствие для поля Тип задания по техническому обслуживанию и т. д. ( Торговля , затем Вариант задания по техническому обслуживанию , затем Тип задания по техническому обслуживанию , затем Актив , затем Модель , затем Производитель , а затем Тип актива ).Если совпадений не найдено, используется запись по умолчанию, в которой выбран только тип задания обслуживания.
Идентификатор действия проекта автоматически связывается с каждой строкой по умолчанию типа задания обслуживания, которую вы создаете. Деятельность по проекту создается в прогнозируемом проекте, выбранном в поле Проект прогноза технического обслуживания на вкладке Активы на странице Параметры управления активами . Целью проектной деятельности является управление прогнозами по часам, предметам и расходам в отношении рабочих заданий.Прогнозы типа задания обслуживания автоматически переносятся в строку заказа на работу и копируются из проекта прогноза в проект заказа на работу, созданный для строки заказа на работу. Целью проектной деятельности является управление прогнозами в отношении рабочих заданий.
Можно настроить пакетное задание для обновления ссылок по умолчанию для типа задания обслуживания через регулярные промежутки времени или запустить задание вручную. Чтобы создать пакетное задание или выполнить обновление вручную, выберите Управление активами > Периодическое > Профилактическое обслуживание > Обновление ссылок по умолчанию для типа задания обслуживания .
Обзор типов работ по техническому обслуживанию, связанных с активами
После создания требуемых комбинаций типов заданий обслуживания по умолчанию можно использовать страницу Все активы , чтобы получить обзор текущего типа задания обслуживания по умолчанию, связанного с конкретным активом. В обзоре показаны все комбинации типов заданий обслуживания по умолчанию, которые можно использовать для типа актива, выбранного для актива. Эти комбинации включают в себя комбинации, которые имеют вариации вариантов типов работ по техническому обслуживанию и сделок по техническому обслуживанию.
Выбрать Управление активами > Общий > Активы > Все активы или Активные активы .
В списке выберите актив, чтобы просмотреть обзор комбинаций типов работ по обслуживанию.
В области действий на вкладке Общие в группе Сопутствующая информация выберите Типы заданий обслуживания .
На левой панели страницы Типы заданий обслуживания активов отображается список всех комбинаций типов заданий обслуживания, связанных с выбранным активом.
Выберите комбинацию типов заданий по обслуживанию, чтобы просмотреть соответствующие настройки для контрольных списков обслуживания, прогнозов и инструментов. В разделе Подробности на экспресс-вкладке Тип задания обслуживания по умолчанию отображается количество связанных контрольных списков обслуживания, прогнозируемых часов, элементов и т. д., связанных с выбранной комбинацией типов заданий обслуживания.
Чтобы просмотреть сведения о выбранном типе заданий обслуживания, выберите Типы заданий обслуживания .
Автоматическое обновление прогнозов типов работ по техническому обслуживанию
В модуле «Управление активами» можно автоматически обновлять любые изменения в прогнозах типов работ по техническому обслуживанию для часовых затрат, стоимости номенклатур и расходов, которые были обновлены в других модулях.